熱線:021-66110819,13564362870
Email:info@vizai.cn
熱線:021-66110819,13564362870
Email:info@vizai.cn
近年來(lái),由于人們對(duì)氣候變化的認(rèn)識(shí)不斷提高,對(duì)海洋碳通量的研究受到重視,但沿海生態(tài)系統(tǒng)仍然是這方面尚未開發(fā)的領(lǐng)域之一。在這項(xiàng)研究中,通過(guò)評(píng)估凈生態(tài)系統(tǒng)生產(chǎn)量和總生態(tài)系統(tǒng)生產(chǎn)量(NEP和GEP)以及CO2交換率(凈生態(tài)系統(tǒng)交換量,NEE),研究了營(yíng)養(yǎng)海岸生態(tài)系統(tǒng)中碳通量的動(dòng)態(tài)。為了估算NEP和GEP,2007年6月20日至7月20日,在韓國(guó)光洋灣沿不同棲息地類型(鰻草和大型藻床、淺層和深層沉積以及深層巖石海岸)測(cè)量了群落產(chǎn)量和呼吸。植被區(qū)的生態(tài)系統(tǒng)產(chǎn)量明顯高于其他生境類型。具體而言,鰻草床具有最高的每日GEP(6.97±0.02 g C m-2 d-1),具有大量生物量和較高的鰻草生產(chǎn)力,而外部大型藻類植被具有最低的GEP(0.97±0.04 g C m-2 d-1)。此外,大型藻類植被由于其最高的P:R比(2.33),顯示出最高的每日NEP(3.31±0.45 g C m-2 d-1)。此外,根據(jù)NEE數(shù)據(jù),鵝草床通過(guò)空氣-海水界面充當(dāng)CO2匯,碳匯率為0.63 mg C m-2 d-1。總的來(lái)說(shuō),我們發(fā)現(xiàn)植被系統(tǒng)(沼澤草和大型藻類床)的生態(tài)系統(tǒng)產(chǎn)量極高,根據(jù)我們的碳通量箱模型概念圖,與非植被系統(tǒng)相比,植被系統(tǒng)所占面積相對(duì)較小。這些結(jié)果表明,植被生態(tài)系統(tǒng)通過(guò)沿海初級(jí)生產(chǎn)力對(duì)無(wú)機(jī)碳的捕獲效率顯著提高。
海洋是全球最大的碳庫(kù),共含有約38000億噸碳,約為大氣碳總量的60倍,約為陸地生物圈和土壤碳總量的10倍(Ciais等人,2013)。此外,地球碳庫(kù)的通量與海洋過(guò)程有關(guān),主要與它們與陸地生態(tài)系統(tǒng)的聯(lián)系有關(guān)。在這些海洋學(xué)過(guò)程中,海洋初級(jí)生產(chǎn)者的有機(jī)碳固定和大氣-海洋界面的碳交換約占人為碳的50%(Sabine et al.2004)。因此,遠(yuǎn)洋生態(tài)系統(tǒng)的初級(jí)生產(chǎn)力占全球碳匯的很大比例,目前正在與陸地生態(tài)系統(tǒng)一起積極研究這一專題。相比之下,對(duì)沿海生態(tài)系統(tǒng)碳研究的關(guān)注較少,可能是因?yàn)樗鼈冎徽伎偤Q竺娣e的一小部分(約7%)(Borges et al.2005)。此外,陸地外來(lái)有機(jī)物質(zhì)的輸入和自養(yǎng)生物(如維管植物、大型藻類、浮游植物和微型底棲生物)的高密度增加了沿海生態(tài)系統(tǒng)的復(fù)雜性,而在公海通常只考慮浮游植物組合。因此,更難全面描述沿海生態(tài)系統(tǒng)中的碳循環(huán),如果我們要更好地理解這些循環(huán),就需要更詳細(xì)地描述對(duì)這些循環(huán)重要的過(guò)程(Mcleod et al.2011)。
光合作用固定的碳可以通過(guò)幾種途徑,其中一種途徑是埋藏在沉積物下面。雖然由于初級(jí)生產(chǎn)力較高,沿海地區(qū)的碳a埋藏率較高,但大多數(shù)以自養(yǎng)生物為主的沿海生態(tài)系統(tǒng)(如河口、紅樹林和鹽沼)通常由于外來(lái)有機(jī)碳超載而成為大氣的凈CO2源(Chen和Borges,2009年,Mcleod等人,2011年)。事實(shí)上,由于光合作用和呼吸的不同來(lái)源、人為輸入的碳和養(yǎng)分以及植被組合的異質(zhì)性,河口和近岸生態(tài)系統(tǒng)的CO2排放范圍特別廣(Borges et al.2005,Laruelle et al.2010)。在這些系統(tǒng)中,水生植被的高生物量可能通過(guò)在光合作用期間將無(wú)機(jī)碳轉(zhuǎn)化為有機(jī)碳,在沿海碳固存中發(fā)揮關(guān)鍵作用(Mcleod et al.2011,F(xiàn)ourqurean et al.2012)。因此,這些棲息地通常被稱為“熱點(diǎn)”,因?yàn)樗鼈兊母咛脊檀媛逝c快速初級(jí)生產(chǎn)相關(guān)。因此,植被海岸生態(tài)系統(tǒng)中的全球碳埋藏預(yù)算估計(jì)約為111 Tg C y-1,約占海洋中埋藏碳總量的一半(Duarte et al.2005),是海洋碳固存總量的重要組成部分。因此,了解自養(yǎng)碳固定率對(duì)于更好地估計(jì)沿海水域的凈碳通量至關(guān)重要。
在與海洋碳通量相關(guān)的重要因素海洋過(guò)程中,CO2氣體交換是與海洋生態(tài)系統(tǒng)碳通量相關(guān)的凈生態(tài)系統(tǒng)交換(NEE)參數(shù)(Lovett et al.2006)。NEE定義了單個(gè)生物體和種群,甚至整個(gè)群落的固定和呼吸CO2,對(duì)于理解沿海碳循環(huán)非常重要。因此,有必要基于大氣-海水界面碳通量參數(shù)的實(shí)際測(cè)量來(lái)研究沿海碳循環(huán),以了解植被在沿海碳循環(huán)中的作用。
盡管碳質(zhì)量平衡很重要,但植被海岸生態(tài)系統(tǒng)中的碳質(zhì)量平衡很難解釋,因?yàn)闃?gòu)建適當(dāng)?shù)奶寄P褪且豁?xiàng)艱巨的工作(Ver等人,1999年)。相比之下,已經(jīng)為公海開發(fā)了許多碳循環(huán)的概念模型(Randerson等人,2002年,Lovett等人,2006年)。這些模型根據(jù)無(wú)機(jī)碳和有機(jī)碳轉(zhuǎn)化過(guò)程中涉及的幾個(gè)生物和化學(xué)參數(shù)來(lái)估算沿海碳循環(huán)。然而,在沿海地區(qū)通過(guò)光合作用合成的有機(jī)碳可以沉入底棲生物,在那里可以被掩埋或從沿海生態(tài)系統(tǒng)運(yùn)輸?shù)浇K颉R虼耍梢酝ㄟ^(guò)測(cè)量與碳固存相關(guān)的組成部分來(lái)估算無(wú)機(jī)碳和有機(jī)碳的預(yù)算,例如生態(tài)系統(tǒng)凈產(chǎn)量(NEP)、外來(lái)碳輸入量、富碳物質(zhì)從海岸的運(yùn)輸以及底棲生物的碳埋率(Duarte et al.2005)。
基于這些觀點(diǎn),我們將重點(diǎn)放在高產(chǎn)沿海生態(tài)系統(tǒng)中的生態(tài)系統(tǒng)生產(chǎn)(NEP和生態(tài)系統(tǒng)總產(chǎn)量[GEP])和NEE上,以估算自養(yǎng)生物驅(qū)動(dòng)的碳通量。在這項(xiàng)研究中,定量估計(jì)了韓國(guó)南部海岸光陽(yáng)灣海岸生態(tài)系統(tǒng)中的GEP、NEP和NEE,其中存在三個(gè)底棲自養(yǎng)生物群落(鰻草、大型藻類和微型底棲生物)和一個(gè)浮游植物群落。具體而言,本研究的目的是1)通過(guò)各種自養(yǎng)生物的光合碳吸收和呼吸來(lái)估算GEP和NEP,2)估算亞生態(tài)系統(tǒng)類型(鰻草床、大型藻類床、未種植的淺層沉積物、深層沉積物和深層巖石棲息地)的NEE,(3)構(gòu)建了當(dāng)?shù)刂脖缓0渡鷳B(tài)系統(tǒng)內(nèi)不同植被類型CO2通量的概念箱模型。
研究地點(diǎn)描述
光洋灣是一個(gè)淺層半封閉系統(tǒng),位于朝鮮半島南部海岸,其水文受通過(guò)狹窄海峽(約4 km寬)與近海水域交換的控制(圖1)。該灣的最大潮差約為4 m。淺海岸地區(qū)(水深小于10 m)占整個(gè)光洋灣地區(qū)的50%以上,主要由淤泥和粘土組成的沉積物構(gòu)成。盡管位于海灣西南角的哈東燃煤電廠和Yulchon工業(yè)綜合體也是碳、養(yǎng)分和污染物的潛在來(lái)源,但陸地外來(lái)碳和養(yǎng)分的主要來(lái)源還是來(lái)自Sumjin河(圖1)。光陽(yáng)灣的自養(yǎng)群落由浮游植物、微型底棲生物、鰻草(Zostera marina L.)和幾種大型藻類組成,所有這些藻類在整個(gè)海灣中分布不均勻(Kim等人,1998年,Kim和Choi,2004年)。
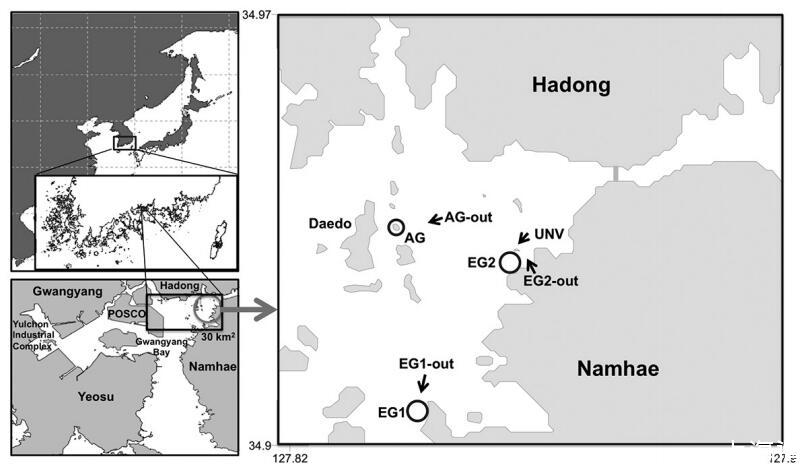
圖1:。研究地點(diǎn)位于韓國(guó)南部海岸光洋灣。子系統(tǒng)通過(guò)自養(yǎng)生物的植被類型和群落組成進(jìn)行區(qū)分,自養(yǎng)生物有植被:AG(大型藻類和浮游植物)和EG(鰻草、浮游植物和微型底棲生物),無(wú)植被:AG out(浮游植物)、EG out(浮游植物和微型底棲生物)和UNV(浮游植物和微型底棲生物)。
在本研究中,根據(jù)初級(jí)生產(chǎn)者的主要類型和基質(zhì)特征,將海灣生態(tài)系統(tǒng)分為五個(gè)棲息地。具體而言,這些是1)鰻草層(EG),2)大型藻類林(AG),3)深層沉積棲息地(UNV;>10 m),4)淺層沉積棲息地(EG out;≤10 m)和5)深巖礁(AG out)(圖1)。利用航拍照片(Daum©、Samah©、Goyang、Korea)的圖像分析和船上調(diào)查相結(jié)合的方法,估算了這五個(gè)棲息地在當(dāng)?shù)叵湫伪壤撸?0 km2)內(nèi)的總面積。這些區(qū)域?yàn)?)鰻草層=1.9 km2,2)大型藻類林=1.6 km2,3)深層沉積棲息地=16.8 km2,4)淺層沉積棲息地=6.3 km2,以及5)深層巖礁=3.4 km2。隨后,選擇了兩個(gè)鰻草床(EG1和EG2),以估算鰻草、浮游植物和微型底棲生物的初級(jí)生產(chǎn)力。選擇一個(gè)大型藻類林分(AG)來(lái)確定每個(gè)大型藻類物種和浮游植物的空間分布,并估計(jì)其個(gè)體初級(jí)生產(chǎn)力。還選擇了四個(gè)非植被化地點(diǎn)、兩個(gè)淺層沉積棲息地(EG1-out和EG2-out)、一個(gè)深部沉積棲息地(UNV)和一個(gè)深部巖礁(AG-out),并估算了常駐自養(yǎng)生物的初級(jí)生產(chǎn)力。最后,我們根據(jù)2007年6月20日至7月20日期間對(duì)自養(yǎng)生物的生產(chǎn)力估計(jì),開發(fā)了海灣碳通量的概念箱模型。
日輻射和環(huán)境因素
使用LI-COR水下光合有效輻射(標(biāo)準(zhǔn)桿數(shù))傳感器(LI-192)在四個(gè)采樣點(diǎn)(EG1、EG2、AG和UNV)的底棲生物附近連續(xù)記錄下涌輻照度,該傳感器連接至數(shù)據(jù)記錄器(LI-1400;LI-COR Inc.,Lincoln,NE,USA),該記錄器在6月20日至21日的原位孵化實(shí)驗(yàn)期間部署,并使用LI-COR傳感器在水面(空氣)上方標(biāo)準(zhǔn)桿數(shù)傳感器(LI-190)。然后使用比爾定律計(jì)算水柱內(nèi)標(biāo)準(zhǔn)桿數(shù)的衰減系數(shù),以找到每個(gè)棲息地中自養(yǎng)生物可用的有效輻照度。在鰻草冠層(約1米深)測(cè)得的輻照度用于計(jì)算鰻草產(chǎn)量,在大型藻類植被最豐富的水域(約4米深)測(cè)得的輻照度用于計(jì)算大型藻類產(chǎn)量。在考慮光衰減系數(shù)的同時(shí),使用空氣輻照度數(shù)據(jù)計(jì)算不同深度的浮游植物產(chǎn)量(Kim等人,2013a)。此外,還利用沉積物表面的輻照度剖面計(jì)算了微型底棲生物的生產(chǎn)力。
在海水采樣期間,測(cè)量了水溫、鹽度和風(fēng)速,并將其用于估算遠(yuǎn)洋生產(chǎn)力和與大氣的CO2氣體交換。具體而言,溫度和鹽度分別使用流浪漢吊墜式溫度記錄器(美國(guó)馬薩諸塞州伯恩市安思特計(jì)算機(jī)公司)和8410-a便攜式鹽度計(jì)(精度±0.003;英國(guó)漢普郡OSIL)進(jìn)行測(cè)量。風(fēng)速數(shù)據(jù)是從距離研究地點(diǎn)約14公里的氣象觀測(cè)中獲得的。
鰻草和大型藻類生物量
在13至15個(gè)樣方(0.063-m2)范圍內(nèi)估計(jì)鰻草枝密度,這些樣方由一名戴水肺潛水員隨意放置在每個(gè)鰻草床內(nèi)。使用每個(gè)樣方內(nèi)單個(gè)嫩枝重量的平均值估計(jì)這些樣品的鰻草地上生物量,使用DT-X數(shù)字回聲測(cè)深儀(美國(guó)華盛頓州西雅圖Biosonics Inc.)測(cè)定海灣內(nèi)鰻草床的總面積,并使用高頻傳感器(420 kHz單波束)收集數(shù)據(jù)。回聲測(cè)深儀數(shù)據(jù)顯示,鰻草在研究地點(diǎn)之間均勻分布(Kim等人,2008年)。此外,通過(guò)收集0.063平方米樣方內(nèi)的所有大型藻類來(lái)量化大型藻類物種的生物量,這些樣方沿垂直于AG現(xiàn)場(chǎng)海岸線的三條樣帶線以2米的間隔放置(n=18個(gè)樣方)。采集樣本后,測(cè)量濕重以估計(jì)單個(gè)物種的分布,并根據(jù)不同物種的總和確定樣本中大型藻類的生物量。然后,根據(jù)大型藻類林分的總面積以及每個(gè)物種的質(zhì)量和相對(duì)豐度,估算海灣大型藻類的總生物量。
鰻草和大型藻類的生產(chǎn)力和呼吸
為了估算個(gè)別鰻草和大型藻類的光合作用和呼吸速率,在晴朗的日子(2007年6月20日),在海灣東北部(EG1附近的海灘)進(jìn)行了原位孵化實(shí)驗(yàn)。一名戴水肺的潛水員從每個(gè)棲息地采集了三份鰻草(Zostera marina)和六種最常見的大型藻類的重復(fù)樣品(n=3),并將其放在不透明的桶中移到孵化點(diǎn),以最大程度地減少光照。在手動(dòng)去除附生生物后,選擇了鰻草芽的葉#3(從最年輕的葉開始計(jì)算)和大型藻類的完整葉狀體,在不同光照強(qiáng)度下進(jìn)行原位培養(yǎng),因?yàn)榈谌~的光合作用最接近平均個(gè)體生產(chǎn)力(未公布數(shù)據(jù))。具體而言,培養(yǎng)瓶(約280毫升的細(xì)胞培養(yǎng)瓶;康寧公司,美國(guó)紐約州康寧市)是通過(guò)向每個(gè)培養(yǎng)瓶中注入20μm過(guò)濾的海水(來(lái)自鰻草(EG-2)或大型藻類(AG)收集點(diǎn))并向其接種10μCi的NaH14CO3來(lái)單獨(dú)制備的。然后將瓶子包裹在中性密度屏幕的各層中,以產(chǎn)生七種不同的透光強(qiáng)度(即0、13、17、30、43、67和100%的環(huán)境光)(Kim等人2013a),以構(gòu)建P-i曲線。該系統(tǒng)在水中自然具有浮力,并保持在約10 cm的深度,從而確保穩(wěn)定的環(huán)境溫度,并允許波浪自然攪動(dòng)瓶子,以減少邊界層的形成。該深度的光強(qiáng)度是使用帶有LI-192水下量子傳感器的LI-1400數(shù)據(jù)記錄器測(cè)量的。將每個(gè)樣品培養(yǎng)3小時(shí),這足以測(cè)量氧氣的產(chǎn)生和消耗(未公布的數(shù)據(jù)),同時(shí)最大限度地減少瓶子效應(yīng),如營(yíng)養(yǎng)耗盡和/或樣品活力的退化(Mateo等人,2001年)。在原位培養(yǎng)后,通過(guò)添加4%中性福爾馬林停止每個(gè)瓶子內(nèi)的光合活性。從每個(gè)瓶子中取出小塊14C標(biāo)記的組織(約20 mg),并添加到單個(gè)閃爍小瓶(20 mL)中,以分析新合成的有機(jī)碳的放射性。在這里,首先用0.5 M NaOH和5%Triton X-100在黑暗中溶解植物組織,然后用H2O2氧化,添加10 mL硫酸面粉閃爍雞尾酒(Perkinlemer,Norwalk,CT,USA)。然后使用液體閃爍計(jì)數(shù)器測(cè)定每個(gè)樣品的放射性(Tri-Carb 2100 TR;Packard Instruments Co.,Meriden,CT,USA)(Gómez et al.2007)。隨后,使用Penhale(1977)推導(dǎo)的方程式計(jì)算每個(gè)樣品的碳吸收量。為了計(jì)算碳吸收率,采用海水碳化學(xué)分析的標(biāo)準(zhǔn)方法測(cè)定海水中的總?cè)芙鉄o(wú)機(jī)碳(CT)(Dickson et al.2007)。從這些短期孵化實(shí)驗(yàn)中獲得的結(jié)果測(cè)量了總光合作用(Mateo等人,2001年)。暗呼吸(Rd)是在現(xiàn)場(chǎng)條件下,使用光學(xué)氧傳感器點(diǎn)測(cè)量氧氣消耗率,該點(diǎn)與Fibox 3(PreSens,GmbH,Regensburg,Germany)有光纖連接(Kim et al.2013b)。具體而言,呼吸通過(guò)在黑暗條件下孵育2小時(shí)后的耗氧量計(jì)算,在將氧氣轉(zhuǎn)換為碳單位時(shí)使用呼吸商1.0。
浮游植物的生產(chǎn)力和呼吸
為了估算浮游植物對(duì)生態(tài)系統(tǒng)生產(chǎn)的貢獻(xiàn),利用14C技術(shù)測(cè)定了鰻草層(EG1和EG2)和深巖礁(AG out)的浮游植物初級(jí)產(chǎn)量。海水樣品通過(guò)200μm網(wǎng)格過(guò)濾,以去除大型浮游動(dòng)物和懸浮顆粒。然后將產(chǎn)生的水和浮游植物添加到孵化瓶(約80毫升的細(xì)胞培養(yǎng)瓶;康寧)中,像大型藻類和鰻草一樣,將其包裹在篩網(wǎng)層中。每個(gè)瓶子接種10μCi的NaH14CO3,然后在原地地表水中培養(yǎng)3小時(shí)。培養(yǎng)后,使用極低真空壓力(<50 mm Hg)通過(guò)玻璃膜過(guò)濾器過(guò)濾樣品(Whatman GF/F;英國(guó)白金漢郡GE Healthcare Ltd.)。在分析放射性之前,向每個(gè)過(guò)濾器中添加UltimaGold(Perkinlemer)閃爍雞尾酒(10 mL)。使用液體閃爍計(jì)數(shù)器測(cè)量合成顆粒碳的放射性。然后使用Strickland和Parsons(1972)推導(dǎo)的方程計(jì)算碳吸收。為了計(jì)算碳吸收率,采用海水碳化學(xué)分析的標(biāo)準(zhǔn)方法測(cè)定海水中的CT(Dickson等人,2007年)。考慮到真光帶內(nèi)標(biāo)準(zhǔn)桿數(shù)的衰減系數(shù),通過(guò)整合整個(gè)深度產(chǎn)量計(jì)算浮游植物的面積群落產(chǎn)量(Taylor 1999)。我們從文獻(xiàn)調(diào)查中考慮了水柱中的每日社區(qū)呼吸(CR),因?yàn)檠鯕庀穆蕸]有使用氧氣計(jì)檢測(cè),氧氣計(jì)與電化學(xué)和光學(xué)氧氣探針相連,約占每日總產(chǎn)量的73%(Williams 1998)。
底棲微生物墊的生產(chǎn)力和呼吸
為了估算軟沉積物中微型底棲生物的初級(jí)生產(chǎn)力和呼吸,從植被覆蓋的淺層沉積(EG2-in)、非植被覆蓋的淺層沉積(EG2-out)和非植被覆蓋的深層沉積(UNV)區(qū)域采集了沉積物樣品。在這項(xiàng)工作中,一名戴水肺的潛水員將透明的丙烯酸管(內(nèi)徑5 cm)插入約12 cm深的沉積物中,而不會(huì)干擾表層沉積物。然后將每個(gè)采樣點(diǎn)的五個(gè)完整沉積物巖芯快速移到實(shí)驗(yàn)室(5小時(shí)內(nèi)),在溫度、鹽度和明暗循環(huán)(分別為23°C、S=32和12:12 LD循環(huán))的模擬現(xiàn)場(chǎng)條件下進(jìn)行保存。此外,樣品適應(yīng)日光熒光燈(20μmol光子m-2 s-1)提供的低光照水平。通過(guò)使用克拉克型氧微電極(尖端直徑25μm;OX-25;Unisense,Aarhus,Denmark)獲得的垂直氧微文件測(cè)量光合作用產(chǎn)生的氧氣,并基于暴露于七種不同光照強(qiáng)度20分鐘后的每個(gè)氧微文件計(jì)算微生物墊的凈面積生產(chǎn)和呼吸,由鹵素照明器提供(KL 2500 LCD;德國(guó)美因茨肖特)(Kühl等人,1996年)。基于Fick的一維擴(kuò)散方程第一定律,考慮沿鹽度和溫度的擴(kuò)散系數(shù),將凈群落產(chǎn)量(NCP)和呼吸(CR)計(jì)算為穿過(guò)擴(kuò)散邊界層的向上和向下O2通量之和(J?rgensen和Revsbech 1985),使用光合(1.2)和呼吸商(1.0)將氧基NCP和CR轉(zhuǎn)換為碳基單位。
凈/總?cè)郝浜蜕鷳B(tài)系統(tǒng)生產(chǎn)
通過(guò)使用Platt等人(1980)的抑制模型擬合光響應(yīng)曲線,估算了鰻草、大型藻類和微藻群落的光合參數(shù)(最大光合速率,PB max;光合效率,α;飽和輻照度,Ek)。這些參數(shù)的平均值和每小時(shí)輻照度剖面再次被替換到Platt等人(1980)的數(shù)學(xué)公式中,以計(jì)算社區(qū)生產(chǎn)力(附錄1)。還考慮了植被的面積生物量,以計(jì)算每日的面積群落產(chǎn)量。在鰻草冠層中心(1米深)和大型藻類物種主要分布的水域(3米深)獲得輻照度數(shù)據(jù),以估計(jì)每個(gè)物種的產(chǎn)量。為了估算底棲微生物墊的群落產(chǎn)量,還對(duì)非植被和淺層沉積生境的深度<3 m以及深層沉積生境的深度>3 m進(jìn)行了估算。利用不同深度和每個(gè)地點(diǎn)的輻照度數(shù)據(jù),在24小時(shí)內(nèi)對(duì)這些數(shù)據(jù)進(jìn)行匯總,得出每日群落產(chǎn)量(Kim et al.2013a)。此外,由自養(yǎng)生物驅(qū)動(dòng)的NCP/總社區(qū)生產(chǎn)(GCP)的整體整合代表了NEP/GEP。
海水pCO2和NEE的測(cè)定(CO2交換)
為了確定海水中的碳化學(xué),中午左右從每個(gè)地點(diǎn)采集了表層海水樣本。將樣品快速轉(zhuǎn)移到500 mL Pyrex瓶中,不引入氣泡,然后立即用100μL飽和HgCl2中毒(Dickson等人,2007年)。在VINDTA系統(tǒng)(德國(guó)基爾馬里安達(dá))中,在實(shí)驗(yàn)室使用庫(kù)侖滴定法和電位滴定法測(cè)定總?cè)芙鉄o(wú)機(jī)碳(CT)濃度和總堿度(AT)。每天對(duì)照已知CT和AT值的海水參考材料檢查CT和AT測(cè)量的準(zhǔn)確性和精密度(由美國(guó)加利福尼亞州圣地亞哥斯克里普斯海洋學(xué)研究所A.Dickson認(rèn)證)。CT的測(cè)量精度約為±2μmol kg-1,AT的測(cè)量精度約為±1.5μmol kg-1。海水分壓(pCO2)值是根據(jù)測(cè)量的CT和AT值,使用Mehrbach等人(1973)的碳酸離解常數(shù)計(jì)算得出的。這組熱力學(xué)常數(shù)已證明與實(shí)驗(yàn)室(Lee et al.1996,Millero et al.2006)和現(xiàn)場(chǎng)(Lee et al.2000,Millero et al.2002)對(duì)海洋溫度和鹽度范圍內(nèi)碳參數(shù)的測(cè)量結(jié)果最為一致。使用CO2交換=k×s×(pCO2空氣-pCO2海水)的方程計(jì)算海氣界面的CO2交換,其中k是氣體傳輸速度(cm h-1),s是CO2在海水中的溶解度,pCO2空氣和pCO2海水是空氣和海水之間CO2分壓的差(Weiss 1974)。空氣中的CO2分壓由濟(jì)州島Kosan的年平均值確定(386 ppmv)。氣體傳輸速度計(jì)算為k=0.31×U102×(Sc/660)-0.5,其中U10是10 m高度處的風(fēng)速(m s-1),Sc是給定溫度和水密度下評(píng)估的施密特?cái)?shù)【Sc=2073.1-125.62(溫度)+3.6276×(溫度)2-0.043219×(溫度)(J?hne等人,1987年,Wanninkhof 1992年)。
 相關(guān)新聞
相關(guān)新聞