熱線:021-66110819,13564362870
Email:info@vizai.cn
熱線:021-66110819,13564362870
Email:info@vizai.cn
日輻射和環境因素
在研究期間(2007年6月20日至7月20日),海灣的每日大氣太陽輻照度范圍為3至33摩爾光子m-2 d-1(附錄2A)。根據估計的光衰減系數,EG1和EG2的鰻草冠層深度處的表面輻照度降低至約8-12%,富含生物量的大型藻類站點(AG)處的表面輻照度降低至約15-32%。淺層沉積物(泥草層底部)的輻照度為表面輻照度的5-8%(附錄2B),而在深層未植被沉積場地(UNV),只有1%的表面輻照度到達沉積物表面。相比之下,水溫和鹽度相對一致,分別在20至25°C和S=32.2至32.5之間。研究期間的平均風速為1.27 m s-1。
鰻草和大型藻類生物量
在鰻草床EG1,估計地上鰻草生物量為805.3±138.6 g FW m-2。2007年6月,預計鰻草覆蓋面積和總生物量分別約為15960 m2和13濕噸(表1)。在eelgrass河床EG2,平均地上eelgrass生物量估計為975.0±158.8 g FW m-2,eelgrass覆蓋面積和總生物量估計分別約為52800 m2和51濕噸。
表1:2007年6月,兩個鵝草床的地上生物量、床面積和總生物量
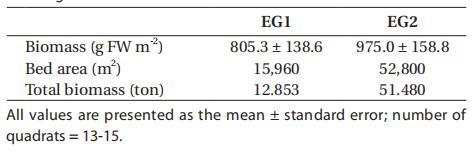
在大型藻類植被現場(AG),沿潮下巖岸分布著11種大型藻類,其中2種為綠藻,3種為褐藻,6種為紅藻(表2)。其中,6個物種(Ulva australis、Gelidium amansii、Gracilaria textorii、Schizymenia dubyi、Chondria crasicaulis和Chondrus ocellatus)占大型藻類總生物量的91%,在4300 m2的大型藻類植被面積中約占2濕噸。一半以上的大型藻類生物量(64%)分布在淺海潮下帶區域(約2米深),81%的生物量分布在水深<4米的區域。在綠藻中,澳大利亞U.australis的個體生物量最高(平均83.12克FW m-2,范圍從0到627.47克FW m-2),占大型藻類總生物量的18%,主要分布在上海岸帶。其中兩種紅藻,克氏絲藻和短尾絲藻的生物量比其他物種高得多,克氏絲藻的生物量介于0到1320.69克FW m-2(平均190.59克FW m-2)之間,短尾絲藻的生物量介于0到1012.66克FW m-2(平均116.19克FW m-2)之間,分別占大型藻類總生物量的41%和25%(表2)。
表2:2007年6月潮下巖岸主要大型藻類種類及其平均生物量
植被(鰻草和大型藻類)和微生物群落(浮游植物和微生物墊)的生產力和呼吸
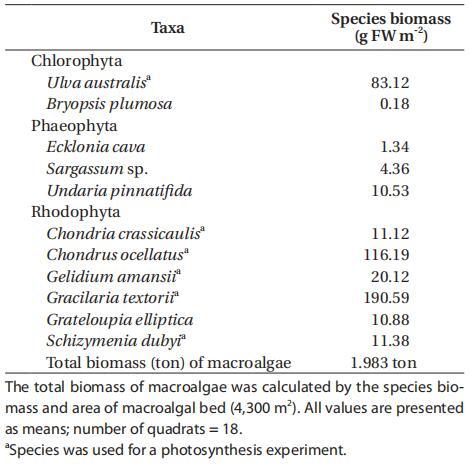
鰻草和大型藻類的光合作用各不相同,P-I曲線的光合參數清楚地確定了其特征(附錄3A-E)。具體而言,鰻草的最大光合速率(PB max)、光合效率(α)和飽和輻照度(Ek)分別為3.28±0.21 mg C g-1 FW h-1、0.008±0.000 mg C g-1 FW h-1/μmol光子m-2 s-1和241±46μmol光子m-2 s-1(表3)。對于大型藻類而言,澳大利亞紫蘇的PB最大值最高(8.31±0.87 mg C g-1 FW h-1),呼吸速率最高(Rd=0.46±0.04 mg C g-1 FW h-1)。五種紅藻(占大型藻類總生物量的78%)的PB最大值均較低,范圍為0.54至2.45 mg C g-1 FW h-1,比澳大利亞紅藻低70-93%。此外,觀察到的最高α值為澳大利亞U.australis(0.032±0.008 mg C g-1 FW h-1/μmol光子m-2 s-1),是其他大型藻類物種的2-10倍。其他紅藻的α和Rd較低,與綠藻和鰻草相當,α的變化范圍為0.003至0.017,而Rd.Z.marina、C.crassicaulis和g.amansii的Ek較高,超過200μmol光子m-2 s-1。鰻草和大多數大型藻類都表現出光抑制的光合特性,除厚殼錦雞兒外,其光照強度大于飽和光照強度。光抑制程度因物種而異,在強光強度下,棉鈴蟲表現出最高的光抑制(總光合作用幾乎得到補償)。
表3:。根據P-I曲線擬合計算的每個物種或群落的光合參數(Platt et al.1980):最大光合速率(PB max)、光合效率(α)、飽和輻照度(Ek)和暗呼吸(Rd)或群落呼吸(CR)
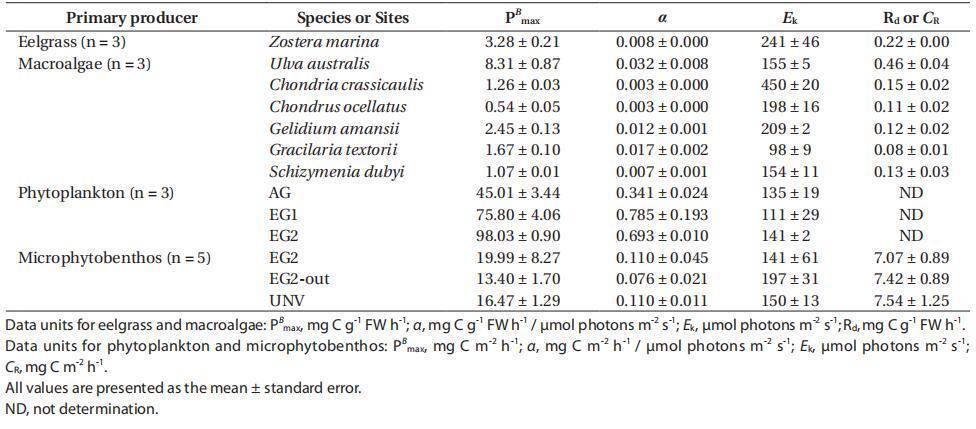
浮游植物的光合特性因植被類型的不同而明顯不同,但微生物群落的光合特性與生境類型的差異不顯著(附錄3H&I)。在鰻草層中觀察到浮游植物的面積PB max和α較高,鰻草層(EG1和EG2)的PB max和α平均比AG高93%,α平均高117%。三個棲息地浮游植物組合的Ek值范圍為111至141μmol光子m-2 s-1,這些值在植被類型之間沒有太大差異。微細胞底棲生物的面積PB max的光合參數在鰻草層最高(EG2,19.99±8.27 mg C m-2 h-1),在鰻草層外側最低(EG2 out,13.40±1.70 mg C m-2 h-1)。鰻草層中微細胞底棲動物的α與深層沉積生境中的α相似(平均值為0.110 mg C m-2 h-1/μmol光子m-2 h-1),該值比鰻草層外的值(EG2 out)高44%。無論棲息地類型如何,微型底棲生物的Ek范圍為141至197μmol光子m-2 s-1。此外,包括在微型底棲生物和微生物呼吸中的沉積物呼吸,在棲息地類型之間沒有顯示出很大的差異,范圍為7.07-7.54 mg C m-2 h-1。總之,優勢大型植物和微生物群落的光合作用和呼吸作用是評估群落和生態系統產量的重要因素。
社區/生態系統生產和NEE
植被群落的GCP較高,但高呼吸速率是植被系統高GCP的潛在抵消因子。具體而言,與微藻群落(浮游植物和微型底棲生物)相比,鰻草類和大型藻類群落表現出極高的NCP和GCP(圖2)。此外,在EG1和EG2中,鵝草床的GCP分別為6.51±0.67和5.70±0.62 g C m-2 d-1,鵝草的NCP分別占EG1和EG2 GCP的35%和10%。大型藻類植被(AG)的NCP占GCP的63%,GCP和NCP值分別為4.83和3.05 g C m-2 d-1。遠洋浮游植物群落是一個凈自養系統(平均值為0.288 g C m-2 d-1),而無論群落類型如何,底棲微生物墊都顯示出凈異養群落(平均值為-0.156 g C m-2 d-1)。
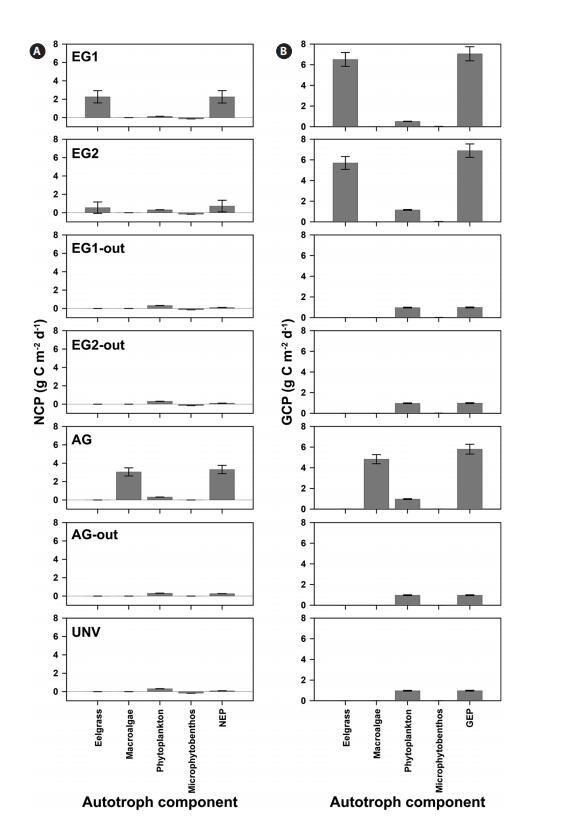
圖2:。2007年6月20日至7月20日(31天),七個地點自養生物的平均每日凈群落生產量(NCP)(A)和每日總群落生產量(GCP)(B)。每日凈生態系統生產量(NEP)和總生態系統生產量(GEP)通過沿自養生物組成的NCP和GCP積分計算。所有值均表示為平均值±標準誤差。
植被系統中GCP和NCP的高比率也導致NEP和GEP值較高(圖2)。根據NEP和P:R比率(GEP與生態系統呼吸,Re),所有研究地點均顯示凈自養生態系統,NEP為0.09-3.31 g C m-2 d-1,P:R比率為1.10-2.33(表4,圖2)。由于鰻草的生產力和生物量較高,因此,鰻草棲息地的GEP最高(EG1,7.06±0.68 g C m-2 d-1)。否則,在未植被覆蓋的巖石海岸(AG out,0.97±0.04 g C m-2 d-1)觀察到最低的GEP,其中僅考慮了浮游植物產量。銀的NEP最高(3.31±0.45 g C m-2 d-1),P:R比最高(2.33)。此外,與GEP相比,鵝草床的NEP(EG1和EG2中分別為2.26±0.67和0.73±0.63 g C m-2 d-1)降低了約78%,即使這些地點由于生態系統的高呼吸而顯示出最高的GCP。
表4:。每月生態系統總產量:每個研究地點的生態系統呼吸(Re)比率(2007年6月20日至7月20日)

根據海水CO2交換測量(NEE)的結果,具有高GEP和密集eelgrass的eelgrass層(EG1和EG2)充當CO2匯(EG1和EG2分別為-0.81±0.17和-0.45±0.09 mg C m-2 d-1)。此外,盡管NEP低于植被系統中的NEP(-0.34±0.07 mg C m-2 d-1),EG1 out仍充當CO2匯。其他未種植場地(EG1外除外)作為CO2源,其結果范圍為0.20至2.32 mg C m-2 d-1(圖3)。否則,即使該系統以高產區(1.66±0.34 mg C m-2 d-1)為特征,銀也可作為大氣的CO2源。
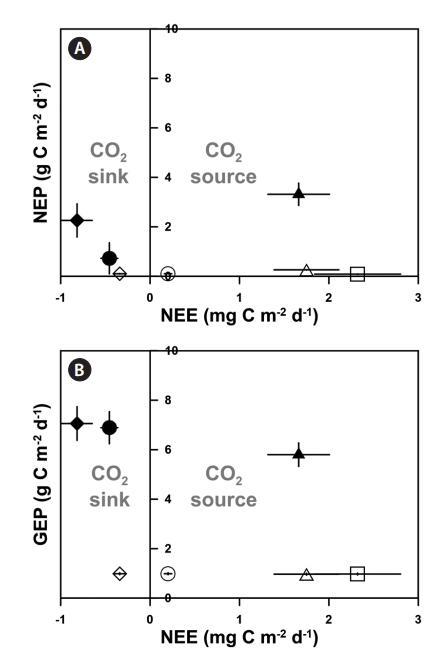
圖3:。兩個鵝草床的凈生態系統產量(NEP)(A)和總生態系統產量(GEP)(B)與海氣界面CO2交換(凈生態系統交換,NEE)的比較(◆,EG1;●,EG2),大型藻床(▲,AG),淺層沉積區(◇,EG1輸出;○,EG2 out),深層沉積區(□),和深部巖石區域(△,2007年6月20日至7月20日(31天)。所有值均表示為平均值±標準誤差。
沿海碳循環的概念模型
沿海碳循環的概念圖,將生態系統生產和NEE沿著各種類型的亞生態系統進行,如圖4所示,在局部方格范圍內(約30 km2)。整個光洋灣系統通過GEP共吸收41.4噸無機碳,并通過Re向沿海水域釋放35.1噸CO2(圖4A)。具體而言,深部沉積區的GEP最高,為18.0噸C d-1。盡管鰻草床只占據了一小部分區域,但與深層沉積棲息地(13.3噸C d-1)相比,該系統占海灣局部箱型規模GEP的73%。大型藻類植被的GEP、NEP和Re最低(分別為0.6、0.3和0.2噸C d-1),可能是因為大型藻類棲息地占據了海灣面積的最小比例。在亞生態系統中,鵝草床的NEP最高(2.8噸C d-1)。因此,這個海灣系統大約有6噸碳,根據NEP可能被固定為有機物,部分光合有機碳可能被隔離到外海。此外,據NEE估計,約47千克CO2從海水排放到大氣中。
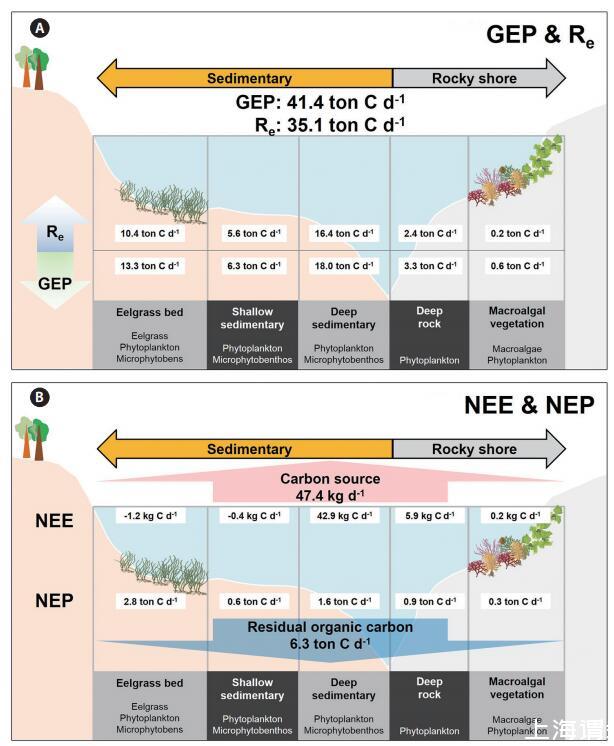
圖4:。光陽灣東北部沿海初級生產者驅動的碳通量概念圖(30 km2)。2007年6月20日至7月20日(31天)期間,不同植被/非植被系統的每日生態系統總產量(GEP)和Re(A)、凈生態系統產量(NEP)和凈生態系統交換(NEE)(B)的平均值。
盡管許多研究試圖描述沿海碳循環,但量化植被沿海生態系統中的碳收支仍然存在局限性,還需要進一步努力(Tokoro et al.2014)。例如,一些研究側重于基于氣體交換和NCP的生態系統生產中的生物代謝和碳質量平衡方法(Bates et al.1998,Sarma 2003)。此外,通過比較河口系統的凈生態系統產量和氣體交換,估算了碳通量(Pradeep Ram et al.2003)。然而,這些研究主要局限于僅考慮浮游植物和/或微型底棲生物的非植被地區。此外,一些研究側重于生物代謝,以估計營養海岸棲息地的碳平衡。因此,通過測量初級生產力,估算了以植被為主的棲息地的碳收支。然而,這些研究沒有考慮許多復雜的碳參數,例如與大氣的氣體交換(Frankignoule和Bouquegneau 1987,Kaldy et al.2002)。為了解決這些不足,我們進行了調查,以建立沿海碳通量的概念箱模型,并關注基于生態系統生產和各種類型植被和非植被系統中CO2氣體交換的碳質量平衡。
沿海植被生態系統通常被描述為高產區,我們的生產/呼吸數據支持這一點。我們研究中的所有亞生態系統在性質上都是自養的(即P:R比率>1)(表4),生產:呼吸比率與其他研究中發現的相當(例如,海草床,1.19-1.50;非植被生境,0.03-34.3)(Duarte和Agusti 1998,Ziegler和Benner 1998)。雖然光陽灣在實驗期間是一個生產力充足的環境,如P:R比率所示,但并非每個亞生態系統都能充當大氣中的CO2匯。這并不奇怪,因為許多具有強烈生物地球化學活動的沿海生態系統可以作為大氣的CO2源(Borges等人,2005年)。我們的NEE結果表明,根據現有植被的類型,植被茂盛的海灣可以同時作為CO2的匯和源(圖3)。具體而言,我們的兩個鰻草床充當CO2匯,可能是因為鰻草的高光合活性和近乎停滯的海水。這些環境特征可能會導致表層海水中pCO2的減少,從而破壞鰻草床內外的海水混合(Madsen et al.2001)。這些特征也發生在我們的非植被地區。具體而言,淺層沉積場地(EG out)的CO2交換速率適中;然而,高能巖石海岸(AG場地)和深海岸沉積場地(AG out和UNV)是大氣的CO2源。與鵝草床相反,由于水平和垂直湍流漩渦和混合,這些棲息地可能導致無機碳從棲息地遷移出去(Madsen et al.2001)。因此,即使大型藻類從周圍海水中吸收大量無機碳,這些棲息地海水中的pCO2可能不會減少。因此,NEE的特點是海岸環境的復雜特征,如海水運動和自養生物生產力減少的陸地有機物流入。
沿海生態系統中的NEP(中上層和底棲NCP的整合)是與向沿海系統以外區域輸出碳相關的一個關鍵因素(Duarte et al.2005),在碳封存方面尤為重要,因為大量輸出的碳可以轉移到公海或埋在沉積物中。尤其是,沉水水生植被顯示出顯著的高生態系統產量(Kaldy et al.2002)。因此,在調節碳循環方面,底棲自養生物的生產是沿海生態系統的一個重要組成部分,它可以提高有機碳固存效率(Mcleod et al.2011)。用五種類型的海岸亞生態系統(鵝草層、大型藻類植被、淺層沉積、深層沉積和深層巖石)劃分的區域NEP代替,構建了當地海岸系統碳通量的概念圖(見圖4)。當地盒狀比例尺(30 km2;光洋灣東北部)的估計NEP值約為6.3噸C d-1,可表示為有機碳質量平衡(圖4A)。特別是,植被棲息地(鰻草和大型藻類床)的NEP在整個研究場地(30 km2)中顯示出33%的GEP,盡管這些棲息地僅占總面積的12%。如果植被覆蓋的沿海生態系統產生超負荷的光合作用有機碳,可能會導致碳固存率增加(Barrón等人,2003年)。有兩條重要途徑可以了解沿海海洋中固定有機碳的去向(Ver等人,1999年)。一部分出口碳將1)運輸到公海(溶解和/或顆粒有機碳),或2)埋在沉積物中。沉積物上層的有機碎屑可能影響營養物質的氧化和循環(Giusti和Marsili Libelli 2005),并通過“大陸架泵機制”出口到邊緣海(Tsunogai et al.1999)。然而,我們的研究沒有通過這兩條途徑對植被海岸生態系統中的有機碳循環進行量化,因此這些因素仍然是未來研究的主題,以澄清海岸有機碳循環。碳排放的另一個次要途徑是排放到大氣中。我們的NEE結果量化了排放到大氣中的CO2,并表明我們當地沿海地區排放到大氣中的CO2約為47.4 kg d-1,約為NEP的200倍。與生態系統生產相比,這一水平很低,但大氣-海水界面的CO2氣體交換尚未就各種植被系統的生態系統生產進行探索。
這項研究確實意義非凡,因為考慮了許多與海岸碳通量有關的可測量參數,如生物量、各種海岸自養生物的生產力以及與原位光剖面的CO2交換。然而,由于與調查相關的困難,本研究未反映時空因素。例如,為了了解植被海灣系統中的沿海碳通量,仍然需要澄清內陸的碳輸入源以及植被現存作物和生產力的異質性,但這些參數受到不同時空尺度的高度影響(Randerson et al.2002)。特別是,NEP的調節高度依賴于初級生產總值和生態系統生產,具有月度時間尺度和區域空間尺度的研究,但河流碳輸出的優先級相對較低。雖然本研究在有限的時空尺度上進行,但應考慮河流碳輸入源,以在一定的時空尺度上完善植物灣系統內的整體概念碳通量模型。此外,目標研究區域的潮汐周期幅度較高,因此,由于光照強度和微藻豐度高度依賴于該因素,因此它對沿海自養生物的生產力有著至關重要的影響。我們不能將復雜的潮汐周期應用于海岸碳循環的概念模型設計,這應該通過未來的研究加以澄清。
總之,本研究提供了一個如何使用質量平衡方法將復雜海岸植被系統中的碳通量估算與光合作用和CO2氣體交換測量相結合的示例。基于我們的結果,我們使用提出的模型對海灣生態系統中的碳通量進行了短期量化。雖然我們的概念箱模型沒有考慮所有復雜的生物活動和環境波動,但它足以評估碳通量的短期和局部尺度估計。此外,我們提出了一個概念性的盒子模型,可應用于其他植被系統,以估算沿海碳循環。為了實現這一目標,需要考慮有關海水中碳化學的其他參數。其中包括溶解無機碳、顆粒/溶解有機碳的去向,以及長期和大空間尺度的水文參數和碳礦化。
確認書
作者感謝Tae Seob Choi和Seung Gu Shin的現場協助和調查。這項工作得到了KIMST/MOF向KYK資助的“造成生態干擾和有害影響的海洋生物管理”項目的支持。
