熱線:021-66110819,13564362870
Email:info@vizai.cn
熱線:021-66110819,13564362870
Email:info@vizai.cn
研究簡介:好氧土壤細菌通過從大氣中清除分子氫(H2)來維持生存。這一關鍵過程是生物地球化學氫循環中的主要匯,支持了貧營養生態系統的生產力。在分枝桿菌屬的M.smegmatis中,大氣H2氧化由兩種系統發育不同的[NiFe]-氫化酶,Huc(2a組)和Hhy(1h組)催化。然而目前尚未解決這些酶如何將從H2氧化中得到的電子傳遞到有氧呼吸鏈的問題。在這項研究工作中,研究人員使用遺傳學方法確認了在氫化酶結構操縱子上編碼的兩種假定鐵硫簇蛋白,HucE和HhyE,對M.smegmatis中的H2消耗是必需的。序列分析顯示,這些蛋白雖然具有同源性,但屬于不同的系統發育分支,并具有不同的金屬結合基序。當編碼這些蛋白的基因被單獨刪除時,H2氧化減少,并且在組合刪除時被消除。相應地,這些缺失菌株的生長產率和長期存活能力與親本菌株相比適度但顯著地降低。在生化和表型實驗中,缺乏假定鐵硫蛋白的突變菌株模擬了氫化酶結構亞基突變株。
假設這些蛋白介導氫化酶的催化亞基和M.smegmatis呼吸鏈中的甲萘醌池之間的電子傳遞;然而其他角色(例如在成熟階段中)也是可能的,需要進一步的工作來解決它們的角色。這些蛋白在大多數編碼Hhy或Huc的有機體中的保守性表明,這些蛋白是大氣H2氧化的重要決定因素。
Unisense微呼吸系統的應用
Unisense微呼吸系統被用來測量和分析M.smegmatis(一種分枝桿菌)在不同遺傳背景下的氫氣(H2)消耗速率。首先在含有甘油的Hartmans de Bont(HdB)最小培養基中培養M.smegmatis的野生型和衍生突變株,直到達到中期靜止相。使用Unisense H2微呼吸微傳感器電極,將其在+800 mV的電壓下極化1小時,并使用已知濃度的H2標準進行校準。在1.1 mL的微呼吸測試瓶中,先后加入靜止相菌液培養物(0.9 mL,OD600=3.0)、H2飽和PBS(0.1 mL)和O2飽和PBS(0.1 mL)。使用Unisense Logger Software記錄H2濃度的變化。當觀察到H2濃度呈線性變化時,計算20秒內H2消耗的速率。
實驗結果
研究確認了HucE和HhyE兩種假定的鐵硫蛋白對于M.smegmatis中H2的消耗至關重要。這些蛋白對于維持氫化酶的活性和細菌的生存能力發揮著必不可少的作用。當HucE和HhyE基因被敲除時,觀察到H2氧化速率顯著降低,表明這兩種蛋白對于氫化酶的功能至關重要。缺乏HucE和HhyE的突變株在生長產率和長期存活能力上表現出降低,這與之前關于氫化酶結構亞基敲除株的研究結果一致。盡管HucE和HhyE在序列上具有一定程度的相似性,但它們在功能上不具有冗余性。在單突變株中沒有觀察到對氫化酶活性的補償現象。HucE和HhyE在大多數編碼Hhy或Huc的有機體中是保守的,這表明這些蛋白在大氣H2氧化中可能具有普遍的重要性。HucE和HhyE可能在氫化酶催化亞基和M.smegmatis呼吸鏈的甲萘醌池之間介導電子傳遞的假設。
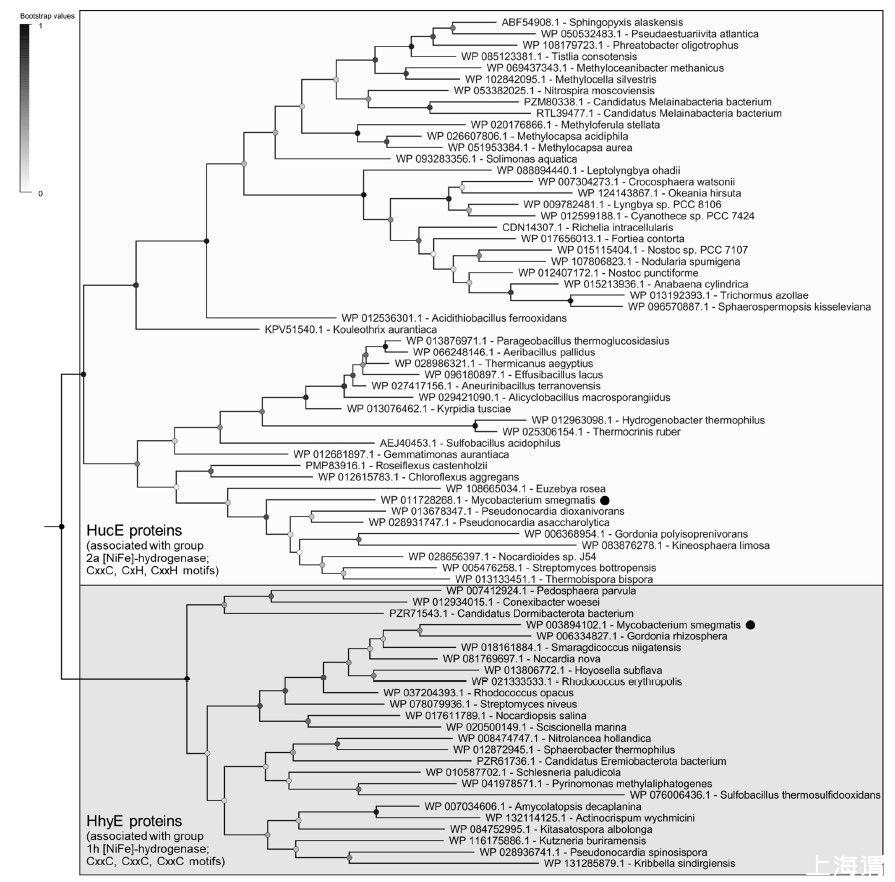
圖1、與2a組和1h[NiFe]-氫化酶相關的HucE和HhyE蛋白的系統發育樹。該樹可視化了52個全長HucE和26個全長HhyE序列的代表性子集之間的進化關系。強調了恥垢分枝桿菌編碼的蛋白質。該樹是使用最大似然法(用部分刪除處理間隙)構建的,用100個重復進行引導,并在中點扎根。
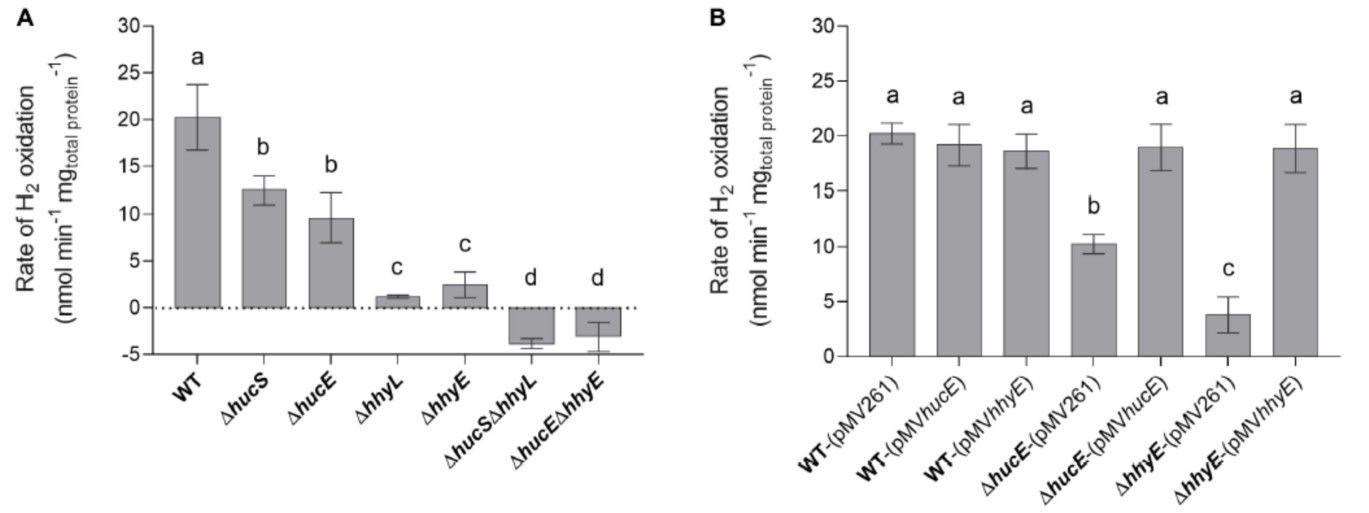
圖2、恥垢分枝桿菌。野生型、衍生突變體和互補突變株的氫氧化。使用Unisense微呼吸H2電極通過電流分析法測量整個細胞在中期穩定期(OD max~3.0后72小時)對H 2的吸收。(A)鐵硫蛋白野生型、單突變體和雙突變體(ΔhucE、ΔhhyE、ΔhucEΔhhyE)以及氫化酶結構亞基單突變體和雙突變體(ΔhhyE)之間H 2氧化速率的比較hucS,ΔhhyL,ΔhucSΔhhyL)。(B)分別用hucE和hhyE的表達補充的ΔhucE和ΔhhyE菌株中的H 2氧化速率。對照包括用空載體pMV261轉化的野生型、ΔhucE和ΔhhyE菌株以及用互補載體pMV hucE和pMV hhyE轉化的野生型菌株。
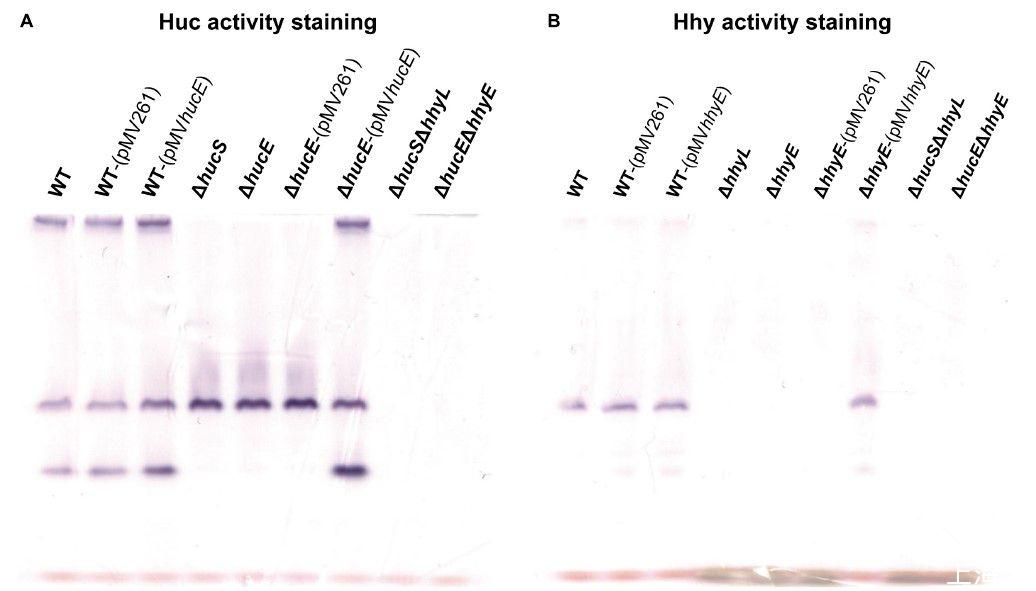
圖3、恥垢分枝桿菌野生型、衍生突變株和互補突變株中的氫化酶活性染色。全細胞裂解物用于在富含H 2的氣氛中以硝基藍四唑作為人工電子受體對H 2攝取進行酶譜染色。(A)野生型、ΔhucS、ΔhucE、ΔhucSΔhhyL、ΔhucEΔhhyE、以及互補的ΔhucE和野生型恥垢分枝桿菌(帶有空pMV261或互補載體pMV hucE)培養物的Huc活性染色)在早期穩定期(OD max~3.0后24小時)收獲。(B)野生型、ΔhhyL、ΔhhyE、ΔhucSΔhhyL、ΔhucEΔhhyE和互補的ΔhhyE和野生型恥垢分枝桿菌(帶有空pMV261或互補載體pMV hhyE)培養物的Hhy活性染色)在中期穩定期(OD max~3.0后72小時)收獲。
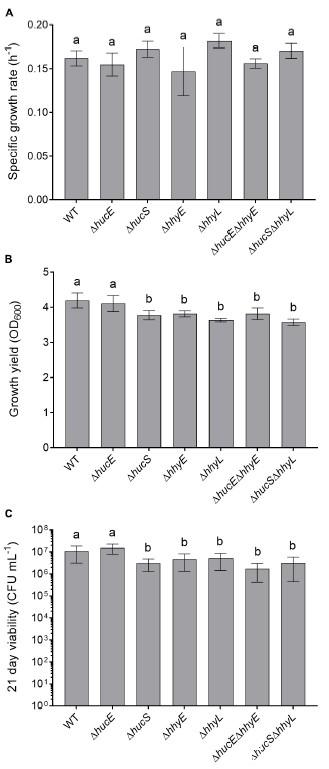
圖4、恥垢分枝桿菌。野生型和突變株的生長和存活率比較。七個菌株在補充有Tween80(LBT)的溶源肉湯中生長:鐵硫蛋白的野生型、單突變體和雙突變體(ΔhucE、ΔhhyE、ΔhucEΔhhyE),以及氫化酶結構亞基的單突變體和雙突變體ΔhucS,ΔhhyL,ΔhucSΔhhyL)。(A)指數期的特定生長率(μ)。(B)穩定期后24小時的最終生長產量(OD max)。(C)穩定期后21天的長期存活率(CFUmL–1)。
圖5、編碼氫化酶的微生物中HucE和HhyE蛋白的分布。(a)編碼2a組[NiFe]-氫化酶的細菌基因組中HucE蛋白的分布。(b)編碼組1h[NiFe]-氫化酶的細菌和古細菌基因組中HhyE蛋白的分布。基因組的顏色取決于是否存在全長蛋白質,是否存在截斷的蛋白質,或者蛋白質是否缺失。
結論與展望
本研究主要介紹了分枝桿菌屬(Mycobacterium)中的M.smegmatis如何通過氧化大氣中的分子氫(H2)來維持生存,這一過程對生態系統的生產力和生物地球化學循環具有重要意義。研究著重于兩個關鍵的[NiFe]-氫化酶:Huc和Hhy,以及它們相關的假定鐵硫蛋白HucE和HhyE。
好氧細菌通過有氧呼吸利用大氣中的H2,這是全球氫循環中的主要過程,并且對低碳土壤生態系統的生產力和生物多樣性起著支持作用。盡管已知M.smegmatis通過兩種氫化酶進行H2氧化,但這些酶如何將電子傳遞到有氧呼吸鏈的具體機制尚不清楚。本研究通過遺傳學方法,構建了HucE和HhyE基因的敲除突變株,以探究這些蛋白在H2消耗中的作用。研究發現,單獨刪除這些基因或同時刪除兩者都顯著降低了H2的氧化速率,并且這種降低與氫化酶結構亞基突變株中觀察到的減少相似。此外缺乏這些鐵硫蛋白的突變株在生長產率和長期存活能力上也表現出適度但顯著的降低。研究結果支持了HucE和HhyE在H2氧化中發揮關鍵作用的假設,并且這些蛋白可能在氫化酶催化亞基和M.smegmatis呼吸鏈的甲萘醌池之間起電子傳遞的作用。
盡管這些蛋白在序列上具有相似性,但它們在功能上不具有冗余性,因為單突變株之間沒有觀察到對氫化酶活性的補償。還同步探討了HucE和HhyE在不同細菌中的保守性和它們在進化上的分布,表明這些蛋白在大多數編碼Hhy或Huc的有機體中是保守的,暗示它們在大氣H2氧化中的重要性。
unisense微呼吸測量技術被用于評估野生型和突變株M.smegmatis的氫化酶活性。通過測量細胞在存在氫氣時的呼吸速率,研究者可以確定氫化酶的功能狀態。通過測量氫氣消耗速率,研究者能夠間接評估不同菌株的生長產率和長期存活能力。這是因為氫化酶活性與細菌的生長和生存能力密切相關。unisense微呼吸測量提供了定量的數據,使得研究者能夠精確地計算氫氣消耗速率,并據此評估氫化酶活性和細菌的代謝狀態。
研究的結論強調了進一步工作的必要性,以解析這些蛋白在氫化酶活性中的具體功能,包括它們是否直接與氫化酶相互作用,以及它們在氫化酶成熟過程中可能扮演的角色。這項研究不僅增進了我們對分枝桿菌能量代謝的理解,揭示了鐵硫蛋白HucE和HhyE在分枝桿菌屬中H2代謝和生存中的關鍵作用,而且為開發新的微生物能源策略提供了潛在的靶點。通過對這些關鍵蛋白的功能研究,可以更好地利用細菌的代謝能力,為生物能源生產和環境修復提供新的途徑。
 相關新聞
相關新聞