熱線:021-66110819,13564362870
Email:info@vizai.cn
熱線:021-66110819,13564362870
Email:info@vizai.cn
解鎖腦功能是探究病理性腦網絡功能障礙的必要條件。由于中樞神經系統形態和生化的復雜性,開發具有預測能力的通用模型必須從體外腦網絡工程入手。基于此,本研究實現了一個基于微電極陣列(MEA)的體外腦網絡,通過體外腦網絡對腦功能控制和病理性腦網絡功能障礙進行了研究,并比較了不同類型的神經元簇的動態特性。研究結果顯示,腦起搏神經元在產生腦網絡電生理模式中起著重要作用,并可能與神經可塑性過程有關。這些發現為未來的計算模型發展提供了有關生理和病理條件下腦網絡行為的重要信息。
微電極陣列(MEAs)上的聚合物裝置:使用聚二甲基硅氧烷(PDMS)約束來塑造網絡連通性。本研究實現了兩種類型的PDMS約束:十字形和圓形掩模。第一種由等臂十字形掩模(圖1a)組成,用于將MEA的有源區域劃分為四個部分,實現四個神經網絡系統(4N)。第二種采用圓形掩模實現單神經網絡系統(1N)。兩種類型的掩膜都是通過將PDMS預聚物和固化劑以10:1的比例混合制成,然后對準并可逆地結合到平面微電極陣列(MEAs)上。
實驗動物:采用第18-19天的Sprague-Dawley胚胎大鼠(E18-19)作為實驗對象。用E18-19胚胎的大腦皮層制備大鼠皮層神經元原代培養物。將神經元原代培養物置于培養基中培育,使神經元組織成形態和功能成熟的神經網絡(圖1b),再進行免疫熒光染色。
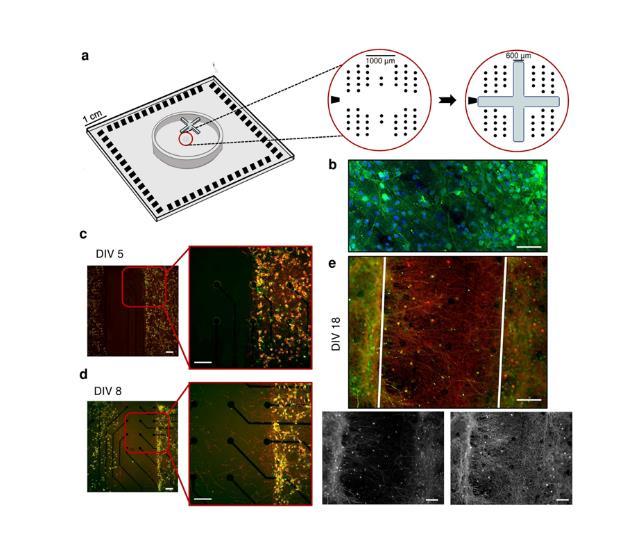
圖1.(a)十字形PDMS掩模在MEA上的定位。黑點表示電極位置;左邊的黑色梯形形狀代表參比電極。MEA的有效區域由1.8 mm×1.4 mm矩形的4個電極簇組成,其中每個電極簇由13個電極組成(?=30μm),另外7個電極位于簇之間。(b)具有代表性的四神經網絡(4N)在體外(DIV)18天的免疫熒光圖像,其中樹突狀微管相關蛋白(MAP2,綠色)和細胞核(DAPI,藍色)被標記。代表性4N培養物的免疫熒光圖像(c)在DIV 5,(d)DIV 8,(e)DIV 18。在兩個不同的神經網絡(簇)之間建立軸突連接,其中樹突微管相關蛋白(MAP2,綠色)和軸突微管相關蛋白(Tau,紅色)被標記。在DIV 5移除十字形掩膜。白色的疊加線劃定了之前被十字形掩模占據的區域。比例尺:100μm。
數據集和實驗方案:本研究中使用的數據集包括16個四網絡系統(4N)和11個單網絡系統(1N)作為對照。為了比較4N和1N組件之間時間序列的激活,將控制網絡的微電極分組為“虛擬集群”,每個微電極由13個單元組成,以保持與4N組件相同的空間組織。此外,為了研究超極化激活的環核苷酸門控(HCN)通道的作用,記錄了3個單網絡系統(1N)的活性,通過計算平均放電速率(MFR)抑制的IC50,得出了HCN抑制劑-伊伐布雷定(IVB)合適濃度的劑量-反應曲線。一旦得出這樣的值(15μM),在1N上進行5次記錄以評估IVB效應。
數據分析:為了檢測脈沖的發生,本研究采用了精確時間脈沖檢測(PTSD)算法。
結果
本研究首先探索并表征了體外互聯亞群(4N)相互作用所產生的自發活動,指出了它們作為起搏器/優勢群體的作用。結果與不顯示模塊化連接的皮質網絡(1N)進行了比較。直到神經元亞群由于PDMS十字形掩膜的存在而保持隔離(圖1a),它們才在每個隔室內建立了密集的連接(圖1b);在去除約束后(DIV 5,體外第5天),種群之間的空間完全沒有任何類型的神經性樹枝(圖1c)。隨后,細胞體開始在裂縫中擴展其樹突,在PDMS移除3天后(DIV 8),神經突束開始向其他亞群擴展,尋找可能的目標(圖1d)。在DIV 18(記錄當天),集群之間的遠程連接結構良好,足夠豐富,可以保證子種群之間的有效連接(圖1e),支持物理模塊化連接的證據。
由模塊化程度調制的峰值和爆發特征
獨立于網絡拓撲類型(即4N或1N),體外皮層網絡表現出自發的電生理活動,其特征是豐富的動態庫,從峰值和爆發信號到通過網絡爆發的網絡全局激活。這種行為與耦合到MEA的同質網絡和集群網絡是一致的。通過比較1N與4N引起的電生理活動,發現在峰值和破裂活動方面存在顯著差異:1N網絡的MFR值高于4N(圖2a),MBR(圖2b)和BD(圖2c)也高于1N網絡。事實上,IBI值呈現相反的趨勢(圖2d)。峰值和爆發統計的宏觀指標由皮質集合的網絡組織調節,而以網絡爆發次數(NB,圖2e)和網絡爆發持續時間(NDB,圖2f)為特征的集體活動沒有顯示出顯著差異。
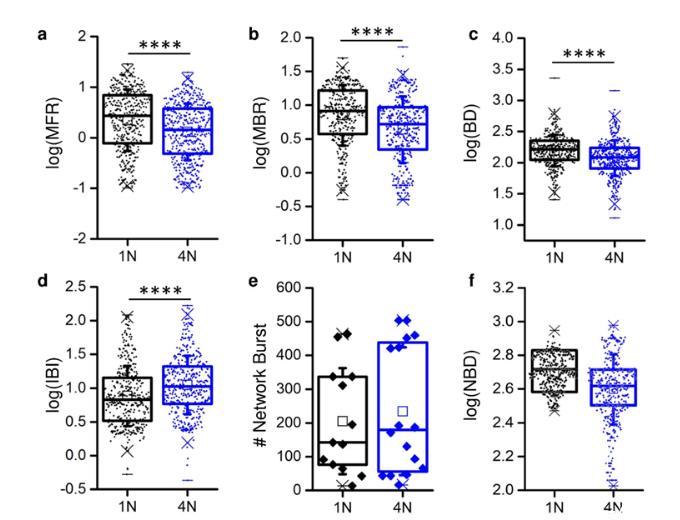
圖2.單網絡(1N,黑色)和四網絡(4N,藍色)組件中的峰值和爆發活動。(a)平均發射速率(MFR),(b)平均爆發率(MBR),(c)爆發持續時間(BD),(d)爆發間隔(IBI),(e)網絡爆發數,(f)網絡爆發持續時間(NBD)。(a-d、f)每個數據點表示相對于單個電極的特定度量(MFR,MBR等)的值。(e)每個數據點相對于多邊環境協定。
由模塊化形成的節律性活動
網絡拓撲并沒有塑造網絡爆發的宏觀度量(圖2e和2f),相反,它們的傳播“模式”受到了影響。為了量化電生理活動的傳播,本研究估計了IFR(瞬時發射速率)。圖3a顯示了1N控制網絡的代表性示例。這四條彩色軌跡代表了用于研究網絡活動的四個“虛擬集群”。在圖3a所示的代表性實驗中,種群事件遵循順時針傳播。通過對集群進行顏色編碼,網絡從紫色集群開始爆發,然后擴散到青色、綠色和紅色集群。另一方面,在圖3b所示的4N中,可以觀察到與在1N中觀察到的相比,簇之間的傳播模式更多變化,更少定型。本研究還表示了三個網絡爆發的IFR曲線:第一個網絡爆發顯示了逆時針傳播,涉及所有四個相互連接的種群(青色-紫紅色-綠色),第二個網絡爆發只涉及三個亞種群(青色缺失),第三個網絡爆發涉及所有隨機序列的群集(既不是順時針也不是逆時針)。
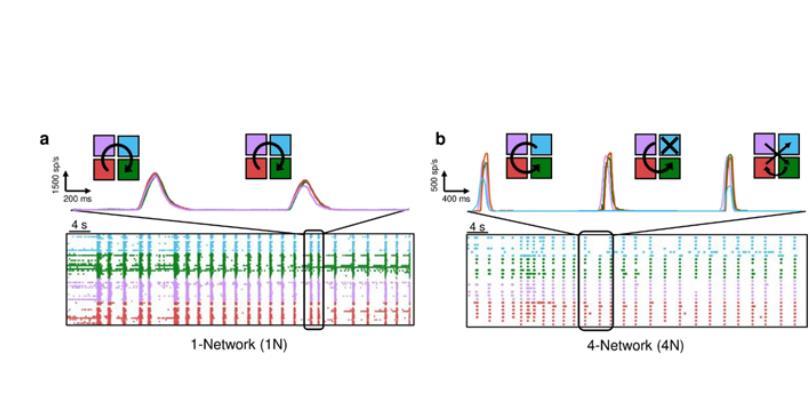
圖3.代表性的(a)單網絡(1N)和(b)四網絡系統(4N)的60 s電生理活動周期。5-s時間窗對應的瞬時發射速率分布在頂部,以及簇激活序列的傳播圖草圖。每種顏色表示一個“虛擬”(1N)或4N集群,而箭頭表示集群之間的網絡爆發傳播。在1N的情況下,涉及到所有的虛擬集群,而在4N的情況下,第一次和第三次網絡爆發涉及到所有的集群,而第二次網絡爆發沒有涉及到青色集群。
探討與結論
本研究使用體外實驗模型探索了腦神經網絡的電生理活動模式,并揭示了起搏神經元活動和神經可塑性在網絡行為中的重要性。通過體外實驗模型,可以對神經網絡進行精細控制和觀察,從而深入研究網絡活動的基本特征。體外模型的實驗設計允許研究人員探索拓撲連接和神經可塑性對網絡活動的影響,為腦神經網絡行為的底層機制提供了重要線索。然而,這項研究是在體外實驗模型中進行的,盡管與體內情況相似,但仍無法完全模擬大腦的復雜性。并且在模擬大腦功能和病理條件時,仍需要更多的研究和實驗來驗證體外模型的準確性和可重復性。
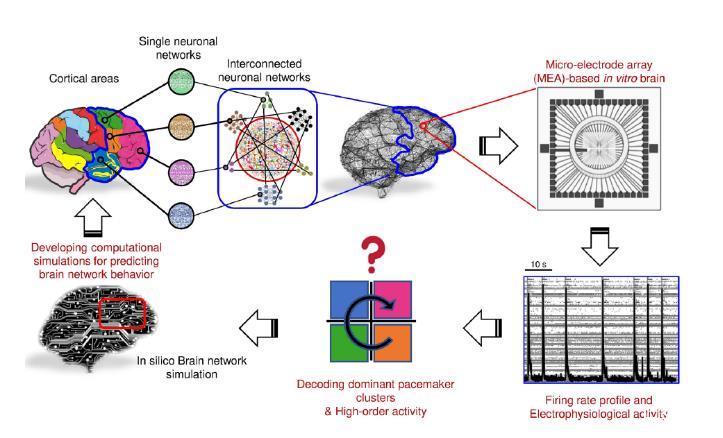
圖4.這項工作的原理和未來展望。主要思想是重新創建與微電極陣列(MEA)耦合的互連神經元網絡,以提取、檢查和定量表征其電生理活動。利用這種方法,我們可以開發出再現所觀察到的神經元網絡活動的計算機模型(數字雙胞胎),從而預測神經網絡的生理行為。從這個角度來看,計算模擬有助于理解神經系統疾病的病因和進展。
總的來說,本研究證實了腦起搏神經元活動和神經可塑性在體外神經網絡的自發活動中的重要作用。實驗結果顯示,腦神經網絡活動具有起搏神經元依賴的節律性和時空固定的模式,可能與神經可塑性有關。這些發現提供了發展計算模型、理解大腦網絡行為和治療神經系統疾病的重要線索。另外,體外模型的應用還可以拓展到轉化醫學研究領域,用于研究神經系統疾病的機制和評估藥物或刺激方案的效果。
