熱線:021-66110819,13564362870
Email:info@vizai.cn
熱線:021-66110819,13564362870
Email:info@vizai.cn
研究簡介:Chloroflexus aggregans是Chloroflexota門(以前稱為Chloroflexi)的代謝多功能、嗜熱、缺氧光養成員,可以光異養、光自養、化能異養和化能自養生長。在溫泉相關的微生物墊中,聚集梭菌與含氧藍藻在動態微環境條件下共存。人們對微堿性、硫化物中房溫泉中的微生物墊的微生物多樣性和功能進行了深入研究。在63–70°C的水溫下,橄欖綠微生物墊以C.aggregans為主,并且未發現產氧藍細菌。在45–62°C的較低溫度下,Chloroflexus spp.與藍藻共存于深藍綠色微生物墊(“藍藻墊”)中。為了闡明C.aggregans的主要生長模式,研究人員研究了中房溫泉微生物墊中晝夜循環中能量代謝和CO2固定相關基因的相對轉錄水平,并與微尺度原位測量相關聯O2和光。宏轉錄組分析表明聚集蛋白具有不同能量代謝模式的兩個時期:中午左右的光養作用和清晨的化能作用。當微生物叢處于高氧狀態(O2 400–800μmolL?1)時,聚集菌主要采用光異養。
Unisense微電極系統的應用
使用Clark型氧微電極(OX-25;Unisense,奧胡斯,丹麥)原位測量O2濃度隨微生物墊深度的變化尖端直徑<25μm、攪拌靈敏度低(<1–2%)和快速響應時間(t 90<0.5 s)。O2微傳感器安裝在電動顯微操作器(Unisense,丹麥)上,并連接到PC接口pA計(Unisense,奧胡斯,丹麥),兩者均由專用數據采集控制、分析和定位軟件(SensorTrace Pro,Unisense,奧胡斯,丹麥)。微操縱器安裝在溫泉旁邊的金屬支架上,允許在自然流動、溫度和光照條件下將微傳感器尖端垂直插入微生物墊中。通過手動操作顯微操作器,將微傳感器尖端小心地定位在墊子表面(定義為0μm)。隨后每15分鐘自動記錄O2微剖面,持續24小時。在每個剖面中,以步徑100μm增量進行O2測量水相并進入生物墊子。對每個深度進行一次測量,并且對于每次測量,應用10秒的等待期,以確保穩定的O2信號,然后記錄O2信號在1S時間內的平均值。
實驗結果
研究表明聚集菌利用其代謝靈活性以及光養和化能生長的能力,在其自然棲息地(中房溫泉微生物墊群落)的不同環境條件下優化其性能。主要的ATP生成時間以及新陳代謝最活躍的時間不僅是中午前后的高光時間(光養性),而且最值得注意的是05:00左右的清晨時間,假設細胞在此時進行化學混合營養生長。利用轉錄組學方法分析了Nakabusa溫泉藍綠色微生物墊中C.aggregans的原位代謝方式。白天光照是主要的能源來源,支持C.aggregans的光自養、光混合自養和光異養生長,而在下午和夜晚,化學自養生長占主導地位。在下午微好氧低光條件下,化學異養生長基于O2呼吸,而在夜晚,在厭氧條件下進行發酵。令人意外的是,在日出前的清晨,C.aggregans細胞似乎進行以O2為終端電子受體的化學自養生長,這表明C.aggregans細胞在墊的微好氧表層進行垂直遷移。

圖1、采樣點和以藍細菌為主的微生物墊的照片。(a)日本中房溫泉“溪流遺址”發育的深藍綠色微生物墊。(b)每個時間點收集的微生物墊核心樣品的直徑為8毫米,面積約為100毫米,15毫米厚。本研究中使用了巖心樣本的上部3毫米。
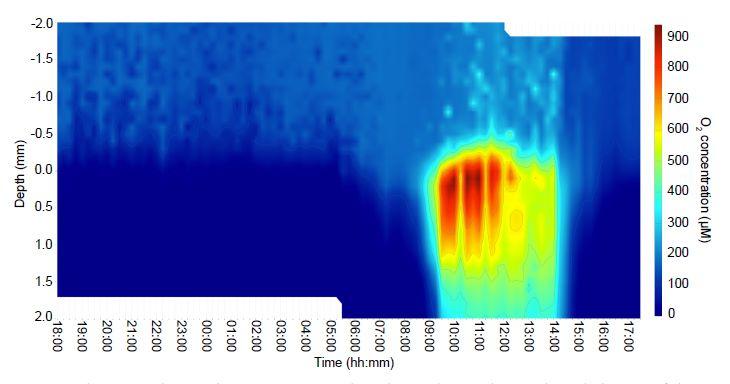
圖2、在晝夜循環中測量的中房溫泉以藍細菌為主的微生物墊中垂直O2濃度分布的熱圖。從11月3日18:00到18日,在24小時內每隔15分鐘測量一次O2濃度(μmol L?1)作為微生物墊深度的函數:11月4日00點。墊子表面用0毫米表示。正深度值表示墊子表面以下的深度,負值表示墊子表面以上的深度,即到溫泉上覆水柱的距離。
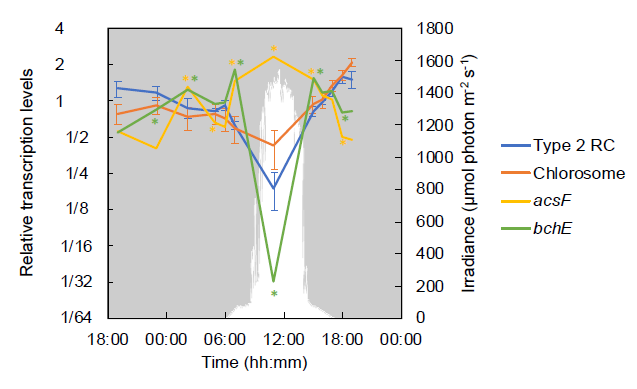
圖3、編碼光合反應中心和葉綠體蛋白的基因以及參與細菌葉綠素生物合成的基因的相對轉錄水平。編碼2型反應中心(RC)(pufLMC:Cagg_1639–1640和Cagg_2631)和葉綠體蛋白(csmAMNOPY:Cagg_1222、Cagg_1209、Cagg_1208、Cagg_2486、Cagg_1206和Cagg_1296)的基因平均值由藍線和橙色線表示。顯示了Mg-原卟啉IX單甲酯需氧環化酶(acsF,Cagg_1285,黃線)和厭氧環化酶(bchE,Cagg_0316,綠線)的值。下降光子輻照度(光合有效輻射[PAR];400–700 nm)以白色表示。星號表示某個時間點顏色對應的特定基因的轉錄與前一個時間點的轉錄顯著不同。
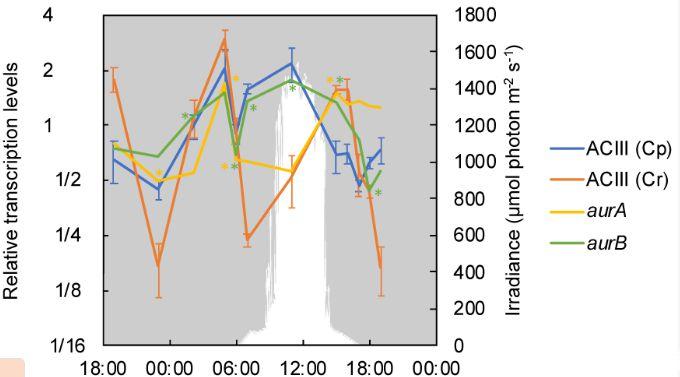
圖4、替代復合物III(ACIII)和金花苷的相對轉錄水平。光養電子轉移的ACIII(Cp、Cagg_3382-3383和3385-3387)和化能電子轉移的ACIII(Cr、Cagg_1523-1527)的平均值分別用藍線和橙色線表示,并帶有標準差。編碼金花苷的aurA和aurB(Cagg_0327和1833)的值分別顯示為黃色(aurA)和綠色(aurB)線。下降光子輻照度(PAR;400–700 nm)以白色表示。星號表示某個時間點顏色對應的特定基因的轉錄與前一個時間點的轉錄顯著不同。
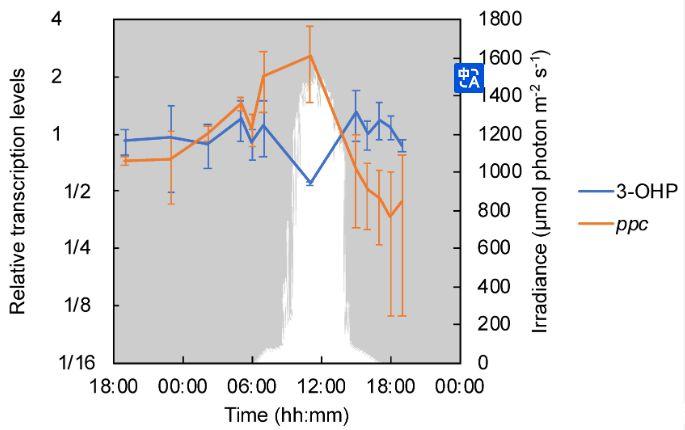
圖5、藍藻墊中編碼3-OHP雙循環關鍵酶和回補途徑相關酶的基因的相對轉錄水平。關鍵3-OHP酶(即丙二酰輔酶A還原酶(Cagg_1256)和丙酰輔酶A合酶(Cagg_3394))以及磷酸烯醇丙酮酸羧化酶(ppc、Cagg_0058和0399)的相對轉錄水平的平均值由藍色表示線和橙色線,分別具有標準偏差。下降光子輻照度(PAR;400–700 nm)以白色表示。
結論與展望
氯光菌聚集體(Chloroflexus aggregans)是一種代謝多樣性、嗜熱、厭氧光營養的葉綠菌門(曾被稱為葉綠菌門)成員,可以以光異養、光自養、化學異養和化學自養的方式生長。在溫泉相關的微生物墊中,氯光菌聚集體在動態微環境條件下與含氧藍藻共存。為了闡明氯光菌聚集體的主導生長模式,研究人員研究了Nakabusa溫泉微生物墊中與一晝夜周期相關的能量代謝和CO2固定相關基因的相對轉錄水平,并與O2和光的微尺度原位測量進行了相關性分析。轉錄組學分析表明,氯光菌聚集體的能量代謝存在兩個不同階段:(1)正午左右的光營養生長和(2)清晨的化學營養生長。在正午時分,當微生物墊富含氧氣(400–800μmol L?1 O2)時,氯光菌聚集體主要采用光異養。在清晨時分,相對轉錄高峰的基因編碼攝取氫酶、碳固定的關鍵酶、呼吸復合物以及TCA循環和乙酸攝取的酶,提示一種好氧的化學混合自養生活方式。這是基于基因轉錄模式的氯光菌聚集體多樣化能量代謝的首次原位研究。在清晨編碼攝取氫化酶、碳固定關鍵酶、呼吸復合物以及TCA循環和乙酸鹽攝取酶的基因的相對轉錄峰值表明有氧化學混合營養生活方式。這是首次基于基因轉錄模式對聚集蛋白的多功能能量代謝進行原位研究。這些研究結果為這些在動態環境條件下茁壯成長的絲狀缺氧光養生物的代謝靈活性提供了新的見解。
 相關新聞
相關新聞