熱線:021-66110819,13564362870
Email:info@vizai.cn
熱線:021-66110819,13564362870
Email:info@vizai.cn
討論
場通量
在該領域,N2Ofluxes 平均為 6.7 6 2 lmol.m-2.h-1,這在其他潮汐研究使用更常見的室通量方法報告的通量范圍內(Middelburg 等人 1995,Wang 等人 2007,Adams et al.2012)。然而,我們的范圍大于潮汐平板文獻中報道的范圍(表 1)。由于方法上的差異,這可能并不令人驚訝。室通量法測量在更大區域內整合的氣體通量(通常為 0.1 m2;Middelburg et al.1995,Wang et al.2007),而微剖面方法基于僅在沉積物中的一個點采集的剖面對氣體通量進行建模。因此,來自微剖面方法的通量被整合到一個小得多的區域,導致更多的變量值。出乎意料的是,幾乎所有的通量都是負的,暴露的沉積物越長,吸收越多(圖 1)。這尤其令人驚訝,因為脈沖事件,特別是潤濕/干燥動力學,通常會導致來自陸地的 N2O 流出量增加 油(Kim 等人,2012 年)。然而,我們調查的外部潮汐脈沖是這些潮灘沉積物中養分可用性的主要驅動因素。因此,我們假設退潮代表了這些沉積物營養物質快速剝奪的時期。
最初,在檢查脈沖潮汐暴露對潮間帶 N2Ofluxes 的影響時,我們預計整個低潮期間沉積物中的氧氣動力學會發生變化。特別是,我們預測,沉積物暴露在大氣中的時間越長,氧氣滲透到沉積物中的深度就越深.反過來,這種增加的氧氣供應可能會刺激硝化作用,從而增加低潮期間 N2O 的產量和隨后的排放量。相反,我們觀察到沉積物氧氣滲透深度隨暴露時間沒有顯著變化(表 1)。這可能是一個結果沉積物含水量(SWC),在整個低潮期間幾乎保持不變,并且總是處于或非常接近飽和狀態(表 1)。這表明,盡管沉積物暴露在空氣中,但沉積物孔隙空間仍然充滿水,并不顯著重新充氧,表明低潮時的 N2Oflux 不是由硝化器的氧氣供應變化驅動的,正如我們最初懷疑的那樣(表 1,圖 1)。
此外,所有產生 N2O 的過程都依賴于向沉積物供應溶解的無機氮,因此,如果 DIN 濃度耗盡,這些過程的速率會減慢,N2O 的產生也會減慢(Burgin 和 Hamilton 2007)。然而,反硝化可以適應通過吸收 N2O 并將其用作終端電子受體來降低底物可用性(Miller 等人,1986 年)。因此,我們假設 N2O 吸收隨沉積物暴露時間顯著增加是由反硝化作用驅動的退潮時微生物群落經歷的營養缺乏(圖 1)。這種 N2O 消耗的增加代表了該系統中脈沖的顯著影響。作為這種營養驅動脈沖的證據,我們可以看看 N2Oflux 和沉積物需氧量。7 月 31 日 N2Oflux 與 SOD 的關系與 8 月 8 日不同;兩者均顯著關系(P,0.05),但對于相同的 SOD, 8 月 8 日的 N2O 吸收量比 7 月 31 日多(P = 0.05)。我們認為這兩個樣本日期表現出對比鮮明的沉積物 N2O 動態,因為它們經歷了非常不同的天氣條件,并且這種天氣變化導致了內部競爭動態的轉變。沉積物。7 月 31 日多霧,有微量降水(NOAA,數據可在線獲取),5 平均沉積物溫度為 24.28C,而 8 月 8 日炎熱晴朗,平均沉積物溫度為 30.58C。這兩天沒有顯著差異,因此,N2Oflux 的差異可能不僅僅是溫度變化的結果。我們假設在陽光明媚的日子里 N2O 吸收增加是由于反硝化器必須與微藻群落競爭用于養分的泥灘;即低潮時的硝酸鹽。這些沉積物中含有豐富的沉積物微藻群落;它們是以光合硅藻為主的組合(稱為微底棲植物 [MPB]),占據了 t 潮灘上只有幾毫米的沉積物(Pascal 等人,2013 年)。作為光合自養生物,MPB 需要包括氮在內的無機養分才能發揮作用,因此不斷與細菌(包括反硝化菌)競爭沉積物中的養分。因此,我們建議多云時,如 7 月 31 日,由于光合速率降低,MPB 養分需求相對較低。這將為包括反硝化菌(產生 N2 或 N2O)在內的細菌留下更多孔隙水養分。然而,在晴天,由于光合作用速率高,反硝化器必須與 MPB 競爭硝酸鹽(Sundback 等人,2000 年)。在這些條件下,孔隙水硝酸鹽可能會更快耗盡,尤其是在退潮時,驅使反硝化器更加依賴 N2O 作為其終端電子受體(Miller et al.1986)。這進一步表明,隨著沉積物潮汐暴露的時間越長,養分的可用性和競爭可能會推動 N2O 的吸收增加,就像 鑒于其消耗 N2O 的能力,N2O 吸收的增加是潮灘沉積物提供的生態系統服務,并且是潮汐淹沒的日常脈沖事件,隨后暴露在大氣中對潮灘氮動力學的顯著影響。
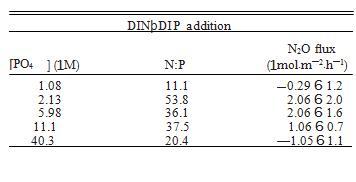
表 2. 每種處理的養分處理濃度、溶解無機氮 (DIN) 與溶解無機磷 (DIN DIP) 的比率,以及每個實驗處理和處理水平下淹沒核心的 N2O 通量。

營養添加實驗
為了區分我們在野外觀察到的 N2Oflux 的特定養分驅動因素,我們在相同潮間帶沉積物的額外核心上設計了一個補充養分添加實驗。這些添加的 N2Ofluxes 的量級通常比我們觀察到的要低得多場(P,0.01)。這可能是沉積物整體代謝率下降的結果,因為 7 月和 8 月的平均沉積物溫度高于 9 月(分別為 27.58C 和 178C)。溫度下降也導致與現場測量的核心相比,養分添加核心中的 SOD 顯著降低(P = 0.01)。因此,我們觀察到的養分添加響應的總體趨勢可能是夏季溫度下發生的情況的保守近似值。
總體而言,我們發現當核心被處理淹沒時 N2Ofluxs 顯著高于核心暴露在空氣中時(P,0.01;圖 2)。這與我們的實地觀察一致,并支持我們的結論,即 N2O 吸收隨著暴露而增加競爭耗盡的養分供應。然而,無論處理如何,對裸露的巖心都沒有顯著的處理效果(圖 2b),表明處理效果是暫時的,只有在沉積物被淹沒時才會持續。一旦暴露,孔隙水養分似乎快速使用,驅使反硝化器越來越依賴間隙和大氣 N2O 進行反硝化,從而導致 N2O 凈負通量進入沉積物(圖 2b)。
3. (a) DIN (y 0.0009x 0.18, R2 0.82, P 0.034) 和 (b) DIN DIP 添加量 (y 0.072x 1.5, R2 0.71, P 0.07) 的淹沒 N2O 通量與 N 與 P 的比率。 箭頭表示處理方向(從低到高的營養添加)。 誤差線表示 6SE,請注意不同的比例。
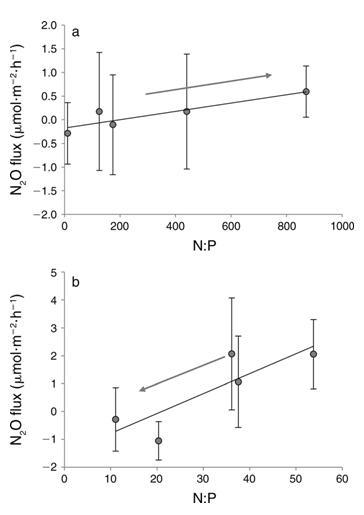
表 2. 擴展。
當核心被處理淹沒時,只有硝酸鹽和 DIN 處理在所有核心中都有顯著的處理效果,盡管 DINtDIP 在所有核心中都是顯著的(圖 2a)。銨處理沒有顯示出處理效果,N2Oflux 與 N2Oflux 之間沒有顯著關系和銨濃度(表 2)。這可能是由于 Michaelis-Menten 動力學。N2O 可以在硝化的第一步產生,氨氧化,這個過程可以通過氨氧化細菌 (AOB) 或氨進行氧化古細菌(AOA)。培養物中的AOA(Candidatus Nitrosopumilus maritimus'strain SCM1)表現出非常低的飽和值(Km);130 nmol/L NH4t,而AOB具有更高的值(Km'100–550 lmol/L NH4t; Martens-Habbena et al.2009。鑒于這些沉積物即使對非常高的銨添加量(超過 600 lmol/L)也沒有表現出任何反應,我們假設這些沉積物中的硝化群落以 AOA 為主,正如已經顯示在一些農業土壤中(Leininger et al.2006。因此,我們認為這些沉積物中的硝化器在非常低的銨濃度下飽和,使它們無法使用額外的底物來提高硝化速率,因此 N2O 的產生是有限的。硝化器缺乏響應添加銨鹽也表明硝化作用不是調節這些沉積物中 N2O 通量的過程,而且信號更可能由反硝化作用和/或 DNRA 支配,如硝酸鹽和 DIN 添加物的顯著處理效果所示(圖.2). 已發現反硝化作用是溫帶(例如,Davidson 等人 1986)和熱帶森林土壤(例如,Bowden 等人,1992 年,Silver 等人,1999 年)以及在一些海洋生態系統中(例如,Dong et al.2002,Wang et al.2007)。
硝酸鹽處理與 N2Oflux 和硝酸鹽濃度之間存在拋物線關系。然而,這種關系完全由 323 處理的 N2Oflux 驅動(5.83 6 3.6 lmol.m—2.h—1),從 0 到 163 的相同關系不是顯著表明沉積物群落對硝酸鹽添加沒有反應,直到它們比環境水平大得多。事實上,除了 323 處理外,所有硝酸鹽添加處理都表現出 N2O 吸收(圖 2a,表 2)。反硝化和 DRNA可以吸收硝酸鹽并產生N2O,盡管每個反應的動力學不同。反硝化在;5至10 lmol/L NO3—飽和(具有Km值),而DNRA在;100至500 lmol/L NO3飽和—(Jorgensen 1989)。鑒于這些 Km 值,我們認為是 DNRA 而不是反硝化,通過產生 N2O 對高水平的硝酸鹽做出反應。這與表明 DNRA 可能在附近鹽沼中勝過反硝化的研究結果一致潮汐小溪 用硝酸鹽施肥(Vieillard 和 Fulweiler 2012)。 從系統中去除哪些生物可利用的氮(例如,Kieskamp et al.1991)。相反,DNRA 的最終產物是銨,氮的另一種生物可利用形式。因此,反硝化有可能抵消增加的 N 負荷,而 DNRA 保持反應性N 在系統內循環。然而,由于動力學,DNRA 可能會在較高的 NO3 濃度下勝過反硝化(Jorgensen 1989)。這表明增加 N 負荷有可能誘導沉積物群落從反硝化向 DNRA 轉變,導致進一步的DIN 水平升高時,生態系統中生物可利用 N 的增加以及沉積物 N2O 排放量可能增加。由于這兩個過程經常在沉積物中同時發生,因此通常很難區分它們對 N2O 通量的單獨貢獻。此外,DNRA 是很少專門測量 N2O 排放;然而,有強有力的證據表明 DNRA 在陸地土壤中的 N2O 排放中起著重要作用,尤其是那些高 pH 值的土壤 (. 6.5) 和高碳含量 (C:N.4;Stevens et al.1998,Morley and Baggs 2010)。
添加 DIN 也有顯著的處理效果,N2Oflux 和 DIN 濃度之間呈正線性相關,表明隨著水柱 DIN 的增加,沉積物 N2O 的產生也增加(圖 2a,表 2)。這遵循類似的線性模式,具有以前在潮間帶沉積物中看到過(Middelburg et al.1995),而不是在河口沉積物中看到的指數關系(Seitzinger and Nixon 1985)。這些發現一起表明沿海生態系統的這兩個組成部分可能對養分負荷有不同的反應。河口沉積物對養分富集的指數響應(Seitzinger 和 Nixon 1985)表明河口 N2O 通量可能比潮間帶沉積物對養分負荷的變化更敏感。已經認為河口沉積物幾乎是環境中 N2O 的唯一來源(Middelburg 等人.1995 年),而對人為 N 的更高敏感性可能只會加劇這種來源(Kroeze 等人,2005 年)。 本研究和其他人(例如,Wang 等人,2007 年)已經表明,dalflats 能夠成為 N2O 的重要匯。由于對 DIN 負載的線性響應,tidalflat N2Ofluxes 可能對 N 的人為增加不太敏感負載比河口高,但增加的 N 負載仍然有可能將這些系統轉移到更持久的 N2O 來源。 2). 令人驚訝的是,這種趨勢與單獨的 DIN 處理相反。我們發現,隨著磷酸鹽濃度的增加,更多的 N2O 被吸收(圖 2a)。這種關系表明,這些沉積物中 N2O 吸收的過程是 P 限制的。 P 限制的進一步證據是 DIN 和 DINtDIP 處理中 N2Oflux 和 N:P 之間的關系(圖 3)。兩種處理都顯示 N2Oflux 和 N:P 之間的正線性相關(圖 3)。然而,斜坡 這些關系的不同之處在于,更重要的是,處理進程對 N2Oflux 具有相反的影響(圖 3)。僅添加 DIN,導致 N:P 增加,產生更多 N2O(圖 3)。 3a). 相反,在 DINtDIP 添加中,DIP 的添加和 N:P 的同時減少,吸收的 N2O 越多(圖 3b)。這些數據一起表明這些沉積物中的 N2O 吸收可能是磷 -勞斯有限。
雖然氮通常被認為是海洋生態系統中的限制性營養物質,但在整個海洋環境中的各種群落中都發現了磷限制(Fourqurean 等人 1992、MacRae 等人 1994、Cotner 等人 1997、Karl 和 Yanagi 1997,Moore et al.2013),并且在來自同一生態系統的潮汐溪沉積物中也發現了 P 限制的證據(Vieillard 和 Fulweiler 2012)。此外,已經發現鹽沼沉積物的異養微生物群落主要受到限制由磷(Sundareshwar 等人,2003 年)。Sundareshwar 等人(2003 年)還表明,當磷限制時 N2O 生成率升高,但在添加磷時降低,這表明反硝化器的限制,正如我們在本研究中觀察到的(圖 3;Sundareshwar 等人 2003。我們的結果表明,這些潮灘沉積物中反硝化器對 N2O 的吸收也可能受到 P 限制。在我們的營養修正處理中,硝酸鹽的存在或不存在,理論上 反硝化 N2O 吸收的驅動因素(Miller 等人,1986 年),不會顯著影響 N2O 吸收。吸收率只會在 P 濃度增加時急劇增加。這表明反硝化和 DIP 之間存在基本且相對未開發的聯系。它還表明與單純的硝酸鹽濃度相比,N:P 的比例對 N2O 吸收的影響可能更大。

圖 1. Rowley River 現場采樣點,2012 年 6 月 29 日。照片來源:AM Vieillard。
該研究表明,較低的 N:P 比率有利于潮灘沉積物中 N2O 的吸收,而較高的 N:P 比率會刺激 N2O 的凈產量(圖 3)。這一發現對沿海生態系統具有潛在的深遠影響。雖然人類活動如肥料生產影響氮和磷循環,在全球范圍內,N 的自然和人為來源比 P 對環境大得多(例如,Fanning 1989,Carpenter 和 Romans 1991,Karl 等人,1997)。因此,人為行為不僅增加了輸送到沿海系統的 N 總量,還增加了 N:P 的比率(Nixon 1993,Benitez-Nelson 2000)。這些對 N:P 比率的擾動在整個生態系統中并不統一,表明受影響更嚴重的生態系統,那些通常具有較高 N:P 比率的生態系統,可能會產生更多的 N2O。 還會抑制潮間帶沉積物從大氣中去除 N2O 的能力,可能導致 N2O 排放量增加。
生態系統 N2O 動態
為了估計我們測量的 N2Ofluxes 對這個系統的潛在影響,我們可以將它們放大到生態系統水平。作為近似值,我們可以使用我們在 6.7 6 2 lmol m-2 h 的實地研究中測量的平均通量—1,并且,假設這個平均值在空間上代表了梅島河口生態系統中 7.2 平方公里的潮汐,我們計算了夏季(6 月、7 月和 8 月),暴露的潮汐為 106 kmol N2O。然后,使用一個范圍在溫帶潮下帶河口沉積物中測量的 N2Oflux 的數量(Seitzinger 和 Nixon 1985,Dong 等人 2002)并將這些值應用于 Plum Island Sound 河口區域,我們可以計算出可能的夏季 N2Ofluxes 范圍約為 26 至 3000 kmol。附近納拉甘西特灣的 N2O 釋放量落在這些值的較低端,當應用于 Plum Island Sound 區域時,可能會從 Plum Island Sound 河口產生 24.5 kmol 的夏季 N2O 排放量。因此,在夏季,潮灘作用作為一個單獨可能更多的水槽 平衡的河口 N2O 產量基于來自附近的大型河口的估計。我們還可以包括來自梅島海峽鹽沼邊緣的 N2Oflux。使用 33 lmol m-2 d-1 的夏季 N2Oflux(Moseman-Valtierra 等人,2011 ) 應用于生態系統中 39 km2 的鹽沼,我們得到了 117 kmol N2O 被沼澤吸收的夏季通量。計算夏季凈生態系統吸收 200 kmol N2O。這是該系統提供的從大氣中去除 N2O 的大量生態系統服務,被歸類為一組被認為是相反的系統(Seitzinger 等人,2007)。 2000,Kroeze 等人,2005 年。此外,這是沿海系統提供的又一項重要服務,受人為活動危害。增加 N 輸送,也許更重要的是,由于人類活動導致沿海系統 N:P 比率增加阻礙的潛力 r 潮汐灘從大氣中去除這種強效溫室氣體的能力。
潮汐脈沖對于溫和潮間帶泥灘中一氧化二氮通量改變的研究——摘要、介紹
潮汐脈沖對于溫和潮間帶泥灘中一氧化二氮通量改變的研究——方法、數據分析
潮汐脈沖對于溫和潮間帶泥灘中一氧化二氮通量改變的研究——結果
 相關新聞
相關新聞