熱線(xiàn):021-66110819,13564362870
Email:info@vizai.cn
熱線(xiàn):021-66110819,13564362870
Email:info@vizai.cn
Cu2+對(duì)NH4+-N、NO3變化的影響?-N、NO2號(hào)?-N、和SOP在一個(gè)周期內(nèi)
圖2a中的數(shù)據(jù)表明,在一個(gè)厭氧-低溶解氧循環(huán)中,對(duì)照反應(yīng)器中NH4+-N的變化與添加Cu2+的反應(yīng)器中的變化幾乎相同,表明Cu2+對(duì)硝化作用沒(méi)有明顯影響(p>0.05)。還發(fā)現(xiàn),在Cu2+濃度為100μg/L時(shí),SOP的平均釋放和吸收分別為54.8和66.1 mg/L,幾乎與對(duì)照試驗(yàn)中觀(guān)察到的結(jié)果相同(54.3和65.4 mg/L)(圖2b)。如圖2所示,兩種NO2?-N和NO3?-在厭氧階段未檢測(cè)到氮,但在低溶解氧階段,其濃度隨時(shí)間增加。在整個(gè)低溶解氧期間,NO3的濃度沒(méi)有顯著差異?-氮在兩個(gè)反應(yīng)器之間(p>0.05),但添加100μg/L的Cu2+會(huì)引起NO2?-氮積累量比對(duì)照低38.9%。Catalan Sakairi等人(1997)還發(fā)現(xiàn),將Cu2+添加到60μg/L的濃度可以最小化NO2?-海水生物脫硝過(guò)程中氮的積累,但其機(jī)理尚未討論。在本研究中,降低NO2的原因?-在接下來(lái)的文本中討論了添加Cu2+的反應(yīng)器中低溶解氧階段的氮積累。
Cu2+對(duì)PHA、糖原和SOP轉(zhuǎn)化及活性污泥生長(zhǎng)的影響
在厭氧-低溶解氧BNR過(guò)程中,在厭氧期間觀(guān)察到SOP釋放、PHA合成和糖原降解,在隨后的低溶解氧階段,PHA被氧化以吸收磷、反硝化、細(xì)胞生長(zhǎng)和補(bǔ)充糖原。從表2可以看出,兩種SBR之間PHA和SOP的轉(zhuǎn)化和細(xì)胞生長(zhǎng)沒(méi)有顯著差異(p<0.05)。顯然,磷累積生物的活性不受Cu2+添加的影響。然而,添加Cu2+導(dǎo)致厭氧或低溶解氧階段的糖原轉(zhuǎn)化比對(duì)照低約25%。
表2添加100μg/L Cu2+對(duì)厭氧-低DO BNR過(guò)程中PHA、SOP和糖原轉(zhuǎn)化以及活性污泥生長(zhǎng)的影響
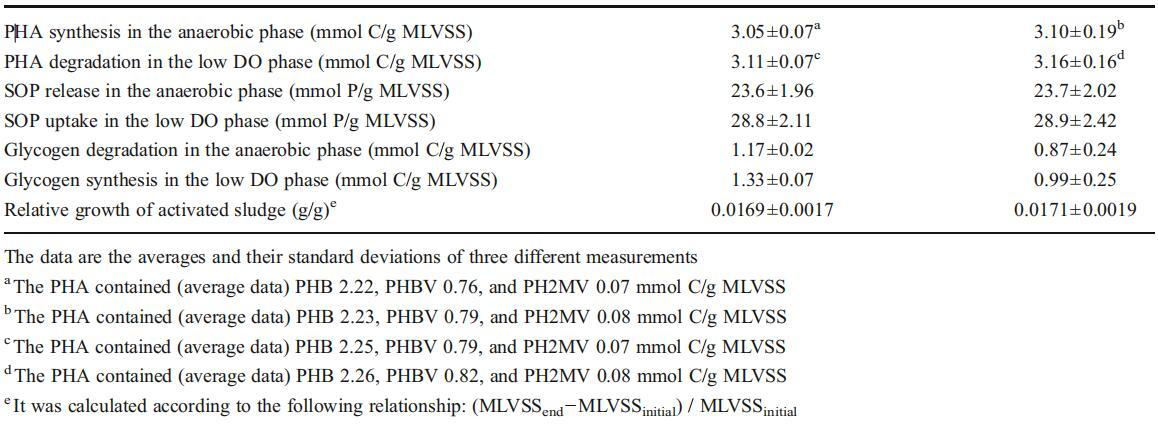
銅離子對(duì)反硝化酶活性的影響
反硝化是一種包含四個(gè)步驟的金屬酶途徑(電子補(bǔ)充材料,圖S8)。NO3號(hào)?還原酶(NAR),NO2?還原酶(NiR)、NO還原酶(NOR)和N2O還原酶(N2OR)是反硝化的四個(gè)關(guān)鍵酶。眾所周知,亞硝酸鹽和N2O的積累分別取決于硝酸鹽和亞硝酸鹽還原以及NO和N2O還原的相對(duì)速率(NO3?→NO2號(hào)?→不→氧化亞氮→N2)。如圖3a所示,與對(duì)照組的0.367 mg N/g MLVSS/min相比,在100μg/L的Cu2+濃度下,NiR活性增加到0.72 mg N/g MLVSS/min,而NAR活性與對(duì)照組相比沒(méi)有顯著影響(0.49 mg N/g MLVSS/min與0.45 mg N/g MLVSS/min)(圖3b)。因此,Cu2+的加入顯著增強(qiáng)了亞硝酸鹽的還原,從而減少了NO2?積累隨著Cu2+的加入,N2OR的活性從0.047(對(duì)照組)增加到0.124 mg N/g MLVSS/min(圖3a),而NOR的活性幾乎與對(duì)照組相同(0.244對(duì)0.264 mg N/g MLVSS/min)(圖3b)。因此,很容易理解,當(dāng)添加100μg/L的Cu2+后,N2OR的活性提高2.63倍時(shí),N2O的生成大大降低。
Cu2+對(duì)活性污泥中反硝化菌群落的影響
nirS、nirK和nosZ基因在編碼反硝化過(guò)程中的關(guān)鍵酶(如亞硝酸鹽還原酶和氧化亞氮還原酶)中起著至關(guān)重要的作用,這些酶可以催化NO2的還原?到NO,最后到N2(Tavares等人,2006年)。因此,根據(jù)文獻(xiàn)(Geets等人2007年;Henry等人2006年),針對(duì)nirS、nirK和nosZ基因的定量實(shí)時(shí)PCR分析可用于確定反硝化細(xì)菌的豐度。圖4說(shuō)明了添加Cu2+后nirS、nirK和nosZ基因拷貝的變化。觀(guān)察到,添加Cu2+后,nirS和nirK基因拷貝數(shù)分別從6.07×106和1.17×106拷貝/g MLVSS增加到6.91×106和1.32×106拷貝/g MLVSS,表明本研究中的反硝化菌數(shù)量通過(guò)添加低劑量的Cu2+顯著增加(p<0.05)。對(duì)照和添加Cu2+的反應(yīng)器中的nosZ拷貝數(shù)分別為7.66×106和9.03×106拷貝/g MLVSS(圖3)。顯然,在添加100μg/L Cu2+后,負(fù)責(zé)N2O還原的反硝化菌數(shù)量顯著增加(p<0.05)。
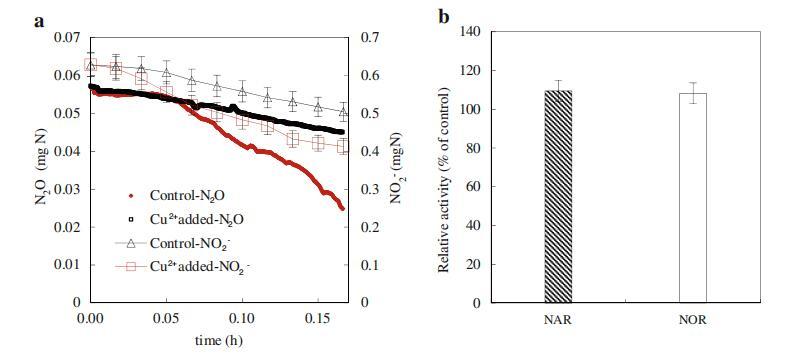
圖3 Cu2+(100μg/L)對(duì)NiR和N2OR(a)以及NAR和NOR(b)活性的影響。誤差條表示三種不同測(cè)量的標(biāo)準(zhǔn)偏差
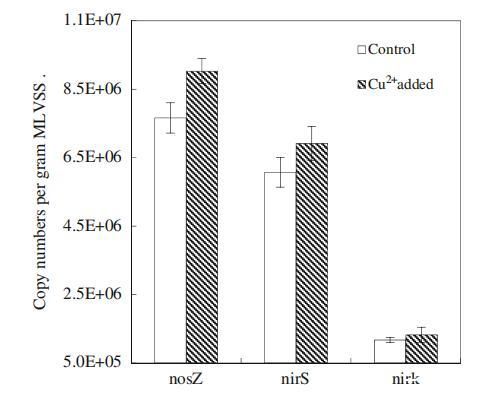
圖4 Cu2+對(duì)nosZ、nirS和nirK基因拷貝的影響。誤差條表示三種不同測(cè)量的標(biāo)準(zhǔn)偏差
另外兩種BNR工藝在城市污水處理過(guò)程中通過(guò)控制Cu2+濃度減少N2O排放的可行性
在另兩種由實(shí)際城市廢水供給的BNR工藝(AAA SBR和MAAA SBR)中,研究了Cu2+對(duì)減少N2O排放的影響。在A(yíng)AA SBR和MAAA SBR中TN和TP的去除效率以及N2O的生成相對(duì)穩(wěn)定后,報(bào)告了數(shù)據(jù)(見(jiàn)表3)。
表3其他兩種BNR SBR中N2O生成和NH4+-N、TN和TP去除效率的比較
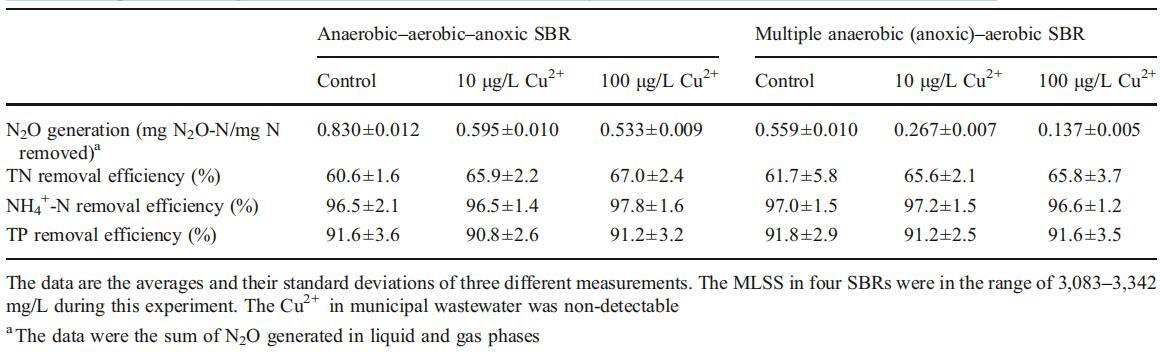
在A(yíng)AA-SBR和MAAA-SBR的對(duì)照反應(yīng)器中,生成的N2O去除率分別為0.830和0.559 mg N/mg N,當(dāng)Cu2+濃度為10μg/L時(shí),去除率分別降至0.595和0.267 mg N/mg N,TN去除率分別從60.6%提高到67%(p<0.05)和61.7%提高到65.8%(p<0.05)。Cu2+從10μg/L增加到100μg/L后,N2O的生成量分別降低到0.533(AAASBR)和0.137 g/L(MAAA-SBR),TN去除效率沒(méi)有顯著影響(p<0.05)。從表3可以看出,與對(duì)照組相比,添加10或100μg/L的Cu2+不會(huì)顯著影響NH4+-N和TP的去除效率。當(dāng)采用厭氧-低溶解氧BNR工藝處理廢水時(shí),所有這些觀(guān)察結(jié)果幾乎與上文討論的結(jié)果相同。因此,可以得出結(jié)論,通過(guò)將廢水Cu2+濃度控制在10至100μg/L,減少城市污水生物處理過(guò)程中N2O的生成是可行的。
據(jù)報(bào)道,反硝化菌stutzeri會(huì)產(chǎn)生缺乏銅的非活性N2O還原酶(Mokhele等人,1987年)。研究還發(fā)現(xiàn),與缺銅培養(yǎng)物相比,增加斯圖澤里梭菌和反硝化副球菌培養(yǎng)物中的Cu2+濃度導(dǎo)致N2O氣體積累減少和生長(zhǎng)加速(Granger和Ward 2003)。從本研究中可以看出,通過(guò)將Cu2+濃度控制在10–100μg/L,不僅可以顯著減少厭氧-低溶解氧生物廢水處理過(guò)程中N2O的生成54.4–72.4%,而且可以提高TN的去除效率,而不會(huì)對(duì)NH4+-N和TP的去除產(chǎn)生不利影響。然而,正如在其他研究中觀(guān)察到的那樣,過(guò)量的Cu2+(5000μg/L)對(duì)BNR微生物有毒(Ganesh等人,2010年;Wu和Rodgers,2010年)。根據(jù)美國(guó)環(huán)境保護(hù)局(2010)的研究,在較低的pH值(<6.5)下,游離銅的毒性水平可能會(huì)抑制硝化作用,在較高的pH值(7–8)下,痕量游離銅(100μg/L是最佳濃度)可能會(huì)加速硝化作用。在本研究中,pH值在7到8之間變化(電子補(bǔ)充材料,圖S9)。因此,當(dāng)前研究中10–100μg/L的Cu2+的存在應(yīng)有利于硝化。然而,未觀(guān)察到Cu2+對(duì)硝化的加速作用,可能是因?yàn)閷?duì)照SBR中的氨氮去除效率高達(dá)97%。
數(shù)據(jù)(圖1和2b)表明,當(dāng)只有少量NO2累積時(shí),SBR中N2O的排放率在有氧期的第一個(gè)小時(shí)內(nèi)較低(低于0.1μmol/min)(SBR#5除外)?-檢測(cè)到N。然后,在接下來(lái)的1.5小時(shí)內(nèi),隨著NO2的積累,N2O的排放速率增加?-N、一些研究人員報(bào)告說(shuō),NO2的積累?-由于亞硝酸鹽或亞硝酸鹽形成的HNO2的毒性抑制了N2O向N2的還原反應(yīng),氮在生物脫氮過(guò)程中誘導(dǎo)了N2O的積累(Itokawa等人,2001年;Zhou等人,2008年)。亞硝酸鹽積累也被認(rèn)為是自養(yǎng)硝化菌反硝化的原因,其最終產(chǎn)物是N2O(Kampschreur等人,2007年)。據(jù)報(bào)道,pH值是影響N2O生成的另一個(gè)重要因素。Th?rn和S?rensson(1996)發(fā)現(xiàn),pH值高于6.8時(shí),缺氧活性污泥中不會(huì)形成N2O。當(dāng)pH值在5到6之間時(shí),N2O產(chǎn)生量最大。在當(dāng)前研究中,添加Cu2+(100μg/L)的SBR在一個(gè)循環(huán)中的pH變化與對(duì)照SBR中的pH變化幾乎相同(電子補(bǔ)充材料,圖S9)。因此,影響N2O排放的因素,包括溶解氧、pH、COD/N、SRT和溫度,在六種SBR中是相同的,這表明NO2的減少?在低溶解氧階段通過(guò)添加Cu2+積累是降低N2O生成的重要原因。
形成亞硝酸銅還原酶催化中心的銅離子已在大量反硝化菌中被報(bào)道(Tavares等人,2006),其耗盡可抑制亞硝酸鹽還原酶的合成(Zumft 1997)。如本研究所示,在活性污泥處理廢水過(guò)程中,添加Cu2+可以顯著提高NiR活性,從而減少NO2?反硝化累積。
在本研究中,觀(guān)察到在污泥濃度(MLSS)約為3200 mg/L的厭氧-低溶解氧BNR工藝處理廢水期間,添加Cu2+可提高N2OR活性,在Cu2+濃度為100μg/L時(shí)觀(guān)察到其最大活性(電子補(bǔ)充材料,圖S10)。N2OR是一種含雙核電子轉(zhuǎn)移位點(diǎn)CuA和催化位點(diǎn)CuZ的銅蛋白(Pomowski等人,2011年)。CuA中心被認(rèn)為接受來(lái)自電子供體(c型細(xì)胞色素和/或假天青蛋白)的電子,并將其轉(zhuǎn)移到N2OR蛋白質(zhì)另一亞單位的相鄰CuZ位點(diǎn),其中N2O結(jié)合并還原為N2和水(Pomowski等人,2011)。銅離子是純反硝化細(xì)菌N2O還原酶生物合成的控制因素(Iwasaki等人,1980年),銅離子缺乏會(huì)導(dǎo)致反硝化污泥的N2OR活性非常低(Miller等人,2009年)。因此,觀(guān)察表明,當(dāng)添加適量的Cu2+時(shí),較高的N2OR活性會(huì)導(dǎo)致BNR過(guò)程產(chǎn)生較少的N2O。
對(duì)活性污泥中nosZ、nirS和nirK基因的實(shí)時(shí)定量分析表明,添加Cu2+促進(jìn)了反硝化菌的生長(zhǎng)。Pieja等人(2011年)報(bào)告稱(chēng),含有甲烷單加氧酶(一種需要銅的酶)的I型甲烷諾菌的生長(zhǎng)可以通過(guò)降低培養(yǎng)基中的Cu2+濃度來(lái)限制。Granger和Ward的研究表明,當(dāng)以N2O為基質(zhì)時(shí),純反硝化細(xì)菌反硝化細(xì)菌在富銅培養(yǎng)基中的生長(zhǎng)速率是在缺銅培養(yǎng)基中的6倍。這些結(jié)果表明,銅離子可能是銅依賴(lài)性細(xì)菌生長(zhǎng)的一個(gè)必要因素。還觀(guān)察到,一些反硝化菌不包含編碼N2O還原酶的基因,例如根癌農(nóng)桿菌C58,這些反硝化菌的最終產(chǎn)物是N2O(Baek等人,2008)。因此,NO2的減少?通過(guò)添加Cu2+積累N2O不僅是由于較高的NIR和N2OR活性,而且還直接受到反硝化菌數(shù)量增加的影響,尤其是能夠還原N2O的反硝化菌。
表2中的數(shù)據(jù)表明,在低溶解氧階段,用于糖原合成的PHA消耗較少,而用于反硝化的PHA節(jié)省較多,這導(dǎo)致添加Cu2+的反應(yīng)器中的TN去除效率高于對(duì)照反應(yīng)器(60.6%對(duì)70.8%)。通過(guò)添加Cu2+改善TN去除的另一個(gè)原因是參與反硝化反應(yīng)的關(guān)鍵酶和微生物增加(圖3)。據(jù)報(bào)道,糖原積累生物(GAO)90%的反硝化最終產(chǎn)物是N2O,這是導(dǎo)致BNR過(guò)程中N2O排放增加的主要原因(Lemaire等人,2006;Meyer等人,2005;Zeng等人,2003)。曾等(2003)指出,在BNR系統(tǒng)中,糖原的轉(zhuǎn)化越少,GAO的活性越弱。因此,結(jié)果表明,銅離子可能會(huì)抑制GAO的活性(見(jiàn)表2),這可能是添加銅的反應(yīng)器中產(chǎn)生較低N2O的另一種可能機(jī)制。
在文獻(xiàn)中,當(dāng)研究生物脫氮除磷時(shí),合成廢水或城市廢水中的Cu2+濃度通常低于10μg/L(Buzier等人,2006;Zeng等人,2003)。根據(jù)本研究結(jié)果,從控制N2O排放和促進(jìn)總氮去除的角度來(lái)看,BNR工藝進(jìn)水中的Cu2+濃度始終不足。此外,本研究發(fā)現(xiàn),約98.6%的進(jìn)水Cu2+被去除,出水中的Cu2+含量低于2μg/L。美國(guó)環(huán)境保護(hù)局(2007)指出,公有處理廠(chǎng)排放濃度為40-200μg/L的Cu2+不應(yīng)對(duì)水生物種構(gòu)成威脅。
在通過(guò)厭氧-低溶解氧工藝處理廢水的過(guò)程中,氮的去除會(huì)經(jīng)歷硝化作用(NH4+-N被氧化為NO2?-N和NO3?-N)和脫氮(NO2?和NO3?還原為NO、N2O和N2),銅的添加顯著影響反硝化作用。此外,本研究表明,銅離子作為一種微量元素,在生物城市污水脫氮除磷過(guò)程的反硝化活性(NiR和N2OR)和群落中起著重要作用,適當(dāng)?shù)你~添加可以顯著減少N2O的生成,同時(shí)提高生物脫氮過(guò)程中的總氮去除率。
致謝
本研究得到了國(guó)家863計(jì)劃(2011AA060903)和污染控制與資源再利用國(guó)家重點(diǎn)實(shí)驗(yàn)室的資助。
 相關(guān)新聞
相關(guān)新聞