熱線:021-66110819,13564362870
Email:info@vizai.cn
熱線:021-66110819,13564362870
Email:info@vizai.cn
鐵作為地殼中豐度最高的氧化還原敏感元素,對湖泊沉積物的氧化還原作用具有重要的指示意義.水生植物根系泌氧在根際形成微域的氧化圈,根際是氧化、還原同時發生的生物活躍區.以輪葉黑藻(Hydrillaverticillata)為研究對象,利用微電極和熒光定量PCR探討根系泌氧作用對沉積物中典型鐵氧化菌(嘉利翁氏菌)和典型鐵還原菌(地桿菌)的影響.結果表明,輪葉黑藻生長迅速,通過根系泌氧作用影響沉積物中鐵的價態和形態,是根際鐵循環的重要參數,并對根際微區微生物有一定的影響.根系泌氧使根際嘉利翁氏菌和地桿菌數量增加,進一步影響根際微生物鐵循環.實驗結果可為微生物對根際鐵循環的研究提供一定的理論基礎。
微生物是湖泊沉積物鐵循環的核心參與者,鐵氧化菌和鐵還原菌在營養元素循環及微量元素的遷移轉化等方面的重要作用,使二者成為目前研究的熱點.本研究選取黑藻為研究對象,研究其根系泌氧作用對沉積物中鐵形態及其對根際沉積物中嘉利翁氏菌和地桿菌的影響,旨在探討植物根系泌氧作用對沉積物中鐵循環的影響。
本研究主要選取浙江省東錢湖(29°52′N,121°34′E)沉積物為研究對象.用彼得森采泥器采集湖中心0~10 cm表層新鮮沉積物,置于便攜式冰箱快速帶回實驗室,鮮泥直接過80目篩(避免沉積物中大顆粒物損壞電極),過篩后混勻,待其自然沉降1 d后備用,實驗開始時沉積物總氮(TN)含量為2.590.14 mg/g,總磷(TP)含量為0.120.03 mg/g,有機質(OM)含量為13.41%1.66%,總鐵(TFe)含量為13.460.44 mg/g,亞鐵(Fe(Ⅱ))含量為6.740.17 mg/g,pH值為7.040.05。
實驗用黑藻采自云南滇池草海,采集后立即帶回實驗室,用自來水沖洗數次并去除里面混雜的水草及其他藻類,然后于實驗室內馴化培養,備用。
將50 ml注射器頂端截掉,用注射器自帶的活塞塞住底部(圖1),加入7 cm左右厚的沉積物,靜置一夜.取性狀一致、長度均為5 cm的黑藻扦插于注射器內,用錫箔紙包裹注射器,保證不透光.對照組不扦插黑藻,其余同實驗組,對照組和實驗組各設置18個重復.放入整理箱(70 cm×50 cm×50 cm)中,小心注入自來水,使水深保持在40 cm左右.放入恒溫室內培養,溫度為25℃培養,12 h/12 h光照/遮光(1500 lx/0 lx).實驗中每天用蒸餾水補足蒸發減少的水分,培養周期為60 d,每10 d取1次樣,取樣方法為對照組和實驗組各隨機取出3支注射器用于后續測定。
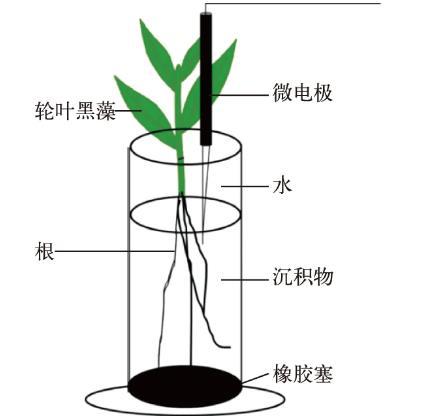
取樣時,小心取出注射器,先用微電極測定植物周圍2 mm處溶解氧的微觀剖面,然后進行破壞性取樣,并將粘附在植物根上的沉積物定義為根際沉積物[18],對照組去除表層的沉積物(1 cm)后定義為非根際沉積物,小心收集后,20℃保存用于理化性質的測定和DNA的提取。
沉積物總鐵(TFe)和亞鐵(Fe(Ⅱ))用3 mol/L HCl提取16 h后,5000轉/min離心5 min,上清液用2,2-聯吡啶分光光度法測定[19].為研究沉積物中鐵氧化物的結晶度以及生物可利用性,采用5步連續提取的方法[11]獲得沉積物中不同形態的鐵.具體做法為:先用1.0 mol/L MgCl2(pH=5)提取可交換態鐵,再用0.1 mol/L焦磷酸鈉提取有機結晶態鐵,然后用0.2 mol/L草酸/草酸銨(pH=3)提取貧結晶態鐵,再用DCB溶液(含0.03 mol/L Na3C6H5O7·2H2O,0.125 mol/L NaHCO3和0.06 mol/L Na2S2O4)提取強結晶態鐵,最后用濃硝酸提取剩余鐵.所有的提取液用2,2-聯吡啶分光光度法測定其中的鐵。
沉積物-水界面的微觀剖面利用微電極進行測量(圖1).實驗用穿刺型氧電極(OXY25,?=25μm,Unisense,丹麥)研究沉積物-水界面幾毫米至幾厘米內的變化.具體做法是:先將微電極連接在四通道主機(Unisense,丹麥)上進行極化和校正.在穿刺樣品時將微電極安裝在1個馬達控制器(MC-232,Unisense,丹麥)上,用顯微操縱器控制(MM 33,Unisense,丹麥)將電極和電腦相連.通過調節相應的參數(響應時間為3 s,步距均為500μm)來研究沉積物-水界面剖面微尺度上的變化.實驗時在黑藻根系周圍2 mm以內進行穿刺(圖1)。
黑藻各項生長指標均隨培養時間的增長而顯著增加,其中,葉片是黑藻進行光合作用的主要場所,隨著培養時間的增加,黑藻生長迅速,生物量不斷增加.黑藻根系隨著培養時間的延長而迅速生長,尤其表現在根系生物量的增加(表2).
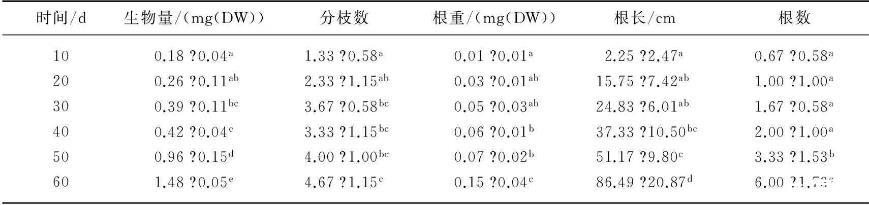
表2不同時間黑藻生長狀況*
*數據為平均值標準差,同列上標不同小寫字母表示差異顯著(P<0.05).
根際微區溶解氧濃度的垂直剖面變化
為了研究種植黑藻后對沉積物-水界面微觀剖面的影響,利用微電極原位研究溶解氧(DO)濃度垂直剖面的變化,結果顯示,黑藻組水體DO濃度為174μmol/L,而對照組僅為137μmol/L(圖2a),這說明黑藻的存在使水體DO濃度增加.當微電極進入泥水界面后,DO濃度迅速下降,至界面下3 mm左右處降為0.當微電極繼續向下到7 mm左右,出現1個明顯的峰值,峰值大小為40μmol/L,峰寬為8 mm,顯著大于黑藻根莖(約1 mm),這進一步說明黑藻根系泌氧的存在。
從有氧層厚度來看,對照組有氧層厚度為5.5 mm左右(圖2b),且隨時間變化不明顯.種植黑藻后有氧層厚度顯著增加(P<0.05),均大于8 mm,且隨培養時間呈增加的趨勢,說明黑藻能使沉積物中DO濃度增加。
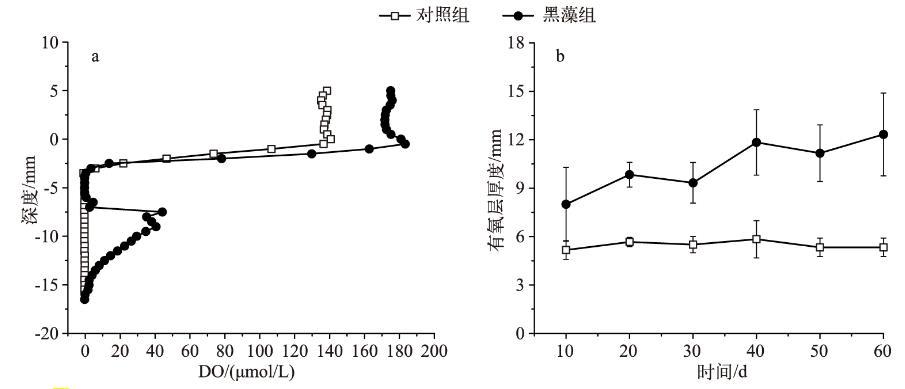
圖2沉積物-水界面DO垂直剖面變化(培養40 d)(a)和有氧層厚度變化(b)
黑藻對沉積物-水界面溶解氧濃度的影響
沉積物-水界面的溶解氧濃度主要受植物根系泌氧和沉積物自身耗氧等因素的影響.本研究利用微電極技術,原位確定了根系泌氧的存在(圖2a)。水生植物根系泌氧是植物在長期淹水缺氧環境中自然選擇的結果,通過植物組織內部強大的通氣組織來實現。黑藻通過葉片進行光合作用產生O2,通過莖、根等通氣組織輸送到植物根系,供根系呼吸、通氣組織是具有巨大空間的海綿組織,它能夠在植物組織內部為氣體存儲和交換提供一個方便的內部通道,使其傳送的氧氣被鄰近組織細胞消耗或擴散到根尖和根際沉積物中[23],并使沉積物-水界面有氧層厚度增加(圖2b)。
結論
1)黑藻的根系泌氧能使沉積物-水界面的溶解氧濃度增加,表層有氧層厚度增加。
2)黑藻通過根系泌氧作用將根際環境中Fe(Ⅱ)氧化,并改變沉積物中鐵的形態,是根際鐵循環的重要參數。
3)黑藻根系泌氧對根際耗氧微區微生物有一定的影響:根系泌氧使根際嘉利翁氏菌和地桿菌數量增加,進一步影響根際微生物鐵循環。
 相關新聞
相關新聞