熱線:021-66110819,13564362870
Email:info@vizai.cn
熱線:021-66110819,13564362870
Email:info@vizai.cn
粉紅色漿果聯合體的系統發育多樣性:16S rRNA基因調查
對粉紅色漿果聚集體中發現的16S rRNA基因的分析證明了一個一致的、簡單的群落,其中兩個系統發育型(定義為97%序列同一性聚類)占測序克隆的65%以上(來自三個樣本的總共273個序列,圖2一種)。PB-PSB1是最豐富的系統發育型(占序列的35-53%),是一種未培養的紫硫細菌(PSB),屬于的Halochromatium-Thiohalocapsa譜系色科(支持信息,圖S1)。
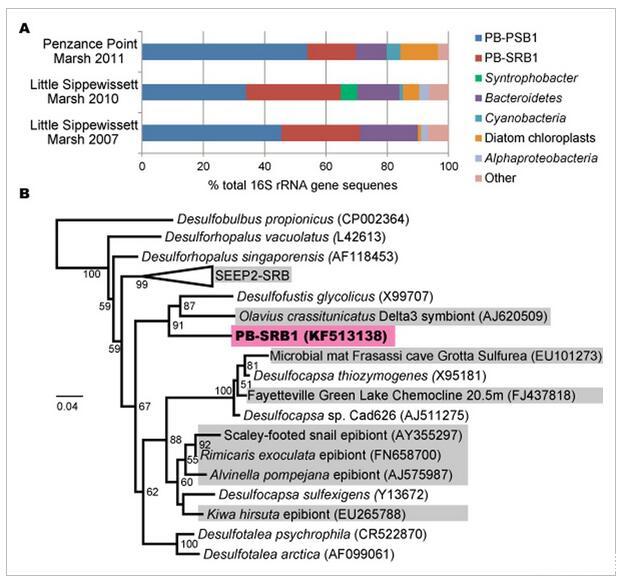
A.16S rRNA基因序列從粉紅色漿果中分離出來,這些漿果采集于2011年(89克隆)彭贊斯角沼澤,2010年(94克隆)和2007年(90克隆)Little Sippewissett沼澤。兩個操作分類單元(OTU),紫硫細菌(PB-PSB1)和硫酸鹽還原菌(PB-SRB1),占觀察到的序列的60%以上。
B.全長PB-SRB1 OTU和相關最大似然系統發育D esulfobulbaceae的。來自未培養生物的環境序列以灰色顯示。超過50%的引導程序支持(500次復制)顯示在節點上。分支長度(和比例尺)對應于各個分支上每個位點的平均核苷酸替換數。
第二豐富的種系型,PB-SRB1(序列15-35%),是最密切相關的Desulfofustis glycolicus,乙醇酸氧化硫酸鹽還原在家庭Desulfobulbaceae(圖2 B)。密切相關的PB-SRB1未經培養的生物體關聯經常發現與參與硫氧化共生或通過硫化物氧化主導的環境的生物體(或化能或光養,圖2的B)。除了硫酸鹽的異化還原外,來自屬的分離物Desulfofustis和Desulfocapsa在外已被證明通過源性硫化物清除劑存在下元素硫和硫代硫酸鹽的歧化而生長(Finster等人,1998年;Finster,2008年)。
其余的16S rRNA基因序列主要是來自海洋硅藻的葉綠體和來自門的不同系統發育型擬桿菌。主要系統發育型擬桿菌的主要屬于來自已知或疑似活性硫循環區域的環境序列進化枝,很少有培養的代表(支持信息,圖S2)。對從粉紅色漿果中擴增的18S rRNA基因序列的分析揭示了與羽狀硅藻和甲藻相關的幾種不同系統發育(支持信息,圖S3)。出于這項工作的目的,我們進一步分析了主要漿果系統發育型PB-PSB1和PB-SRB1。
原位識別與空間布置
粉紅色漿果由透明外聚合物基質中形狀不規則的粉紅色小管組成(圖1 F)。粉紅色漿果薄片的共聚焦顯微鏡顯示,這些小管以紫色硫細菌為主,被鑒定為含有可折射元素硫包裹體的自發熒光球菌(直徑2-4μm)(圖3)。這些折射性內含物在溶劑(甲醇、乙醇)和洗滌劑(SDS、Triton X100)中不穩定,證實了它們是細胞內元素硫球。這些細胞還通過催化報告基因沉積-熒光原位雜交(CARD-FISH)使用GAM42a組特異性探針進行熒光標記,該探針與來自16S rRNA雜交Gammaproteobacteria的(Manz等,1992;支持信息,圖S4)。
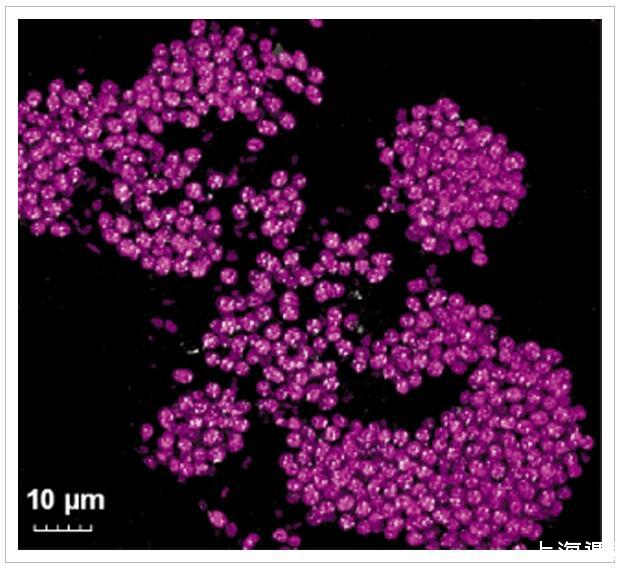
圖3
切片粉紅色漿果組織的表面反射共聚焦顯微鏡。紫色硫細菌的自發熒光(激發543 nm,發射550–570 nm)以粉紅色顯示,來自可折射元素硫包裹體的反射信號以白色顯示。
PB-SRB1物種通過CARD-FISH定位,使用從16S rRNA基因庫設計的系統發育特異性探針(SRB-PiBe213;本研究)。SRB-PiBe213探針與3μm長的桿雜交,這些桿豐富且散布在PB-PSB1細胞的密集島中(圖4)。使用靶向Deltaproteobacteria[DELTA495a-c;Lücker及其同事(2007年)]。與SRB-PiBe213探針雜交的游離細胞很少見(<總細胞數的1%),但可以在潮間帶水池的上覆水中檢測到(數據未顯示)。還發現覆蓋的池水和附近潮汐通道中的粉紅色沙塊含有球狀PSB細胞的微觀聚集體,以及與SRB-PiBe213探針雜交的桿狀細胞。最小的聚集體僅包含5-6個總細胞,而較大的微聚集體的直徑為20-50μm(數據未顯示)。
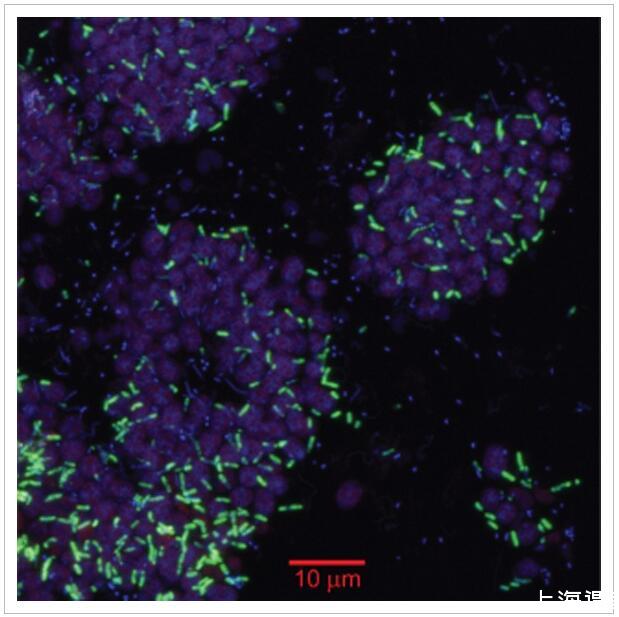
圖4
使用系統發育特異性探針通過催化報告基因沉積-熒光原位雜交(CARD-FISH)鑒定漿果相關的PB-SRB1物種。掃描共聚焦顯微照片顯示了來自聚集體外圍附近的粉紅色漿果生物量的橫截面。該圖像是三個熒光信號的疊加:自發熒光紫硫細菌(以粉紅色顯示,激發543 nm,發射550–570 nm),來自SRB-PiBe213探針的CARD-FISH信號(以綠色顯示,Alexa 488酪胺)和DAPI核酸染色劑(以藍色顯示)。
系統發育多樣性的宏基因組分析
還使用未組裝的Illumina鳥槍宏基因組數據研究了粉紅色漿果中微生物的多樣性,這避免了聚合酶鏈反應(PCR)擴增中固有的偏差。宏基因組數據中不同分類群的豐度和多樣性使用核糖體RNA基因序列(Meyer進行評估et al.,2008)和保守的系統發育標記基因(Darling et al.,2014)。在鳥槍法宏基因組序列中觀察到的多樣性概括了基于PCR的調查(圖5)。一個顯著的例外是存在大量Alphaproteobacteria獵槍數據中,由代表驅動紅細菌目(Oceanicola和Oceanicaulis物種)和紅螺菌目的。根據針對SILVA和核糖體數據庫項目(RDP)數據庫的計算機內特異性搜索(數據未顯示),這種差異可能是由于“通用”8F細菌引物引入的偏差,該引物無法很好地覆蓋這些訂單中的生物體。
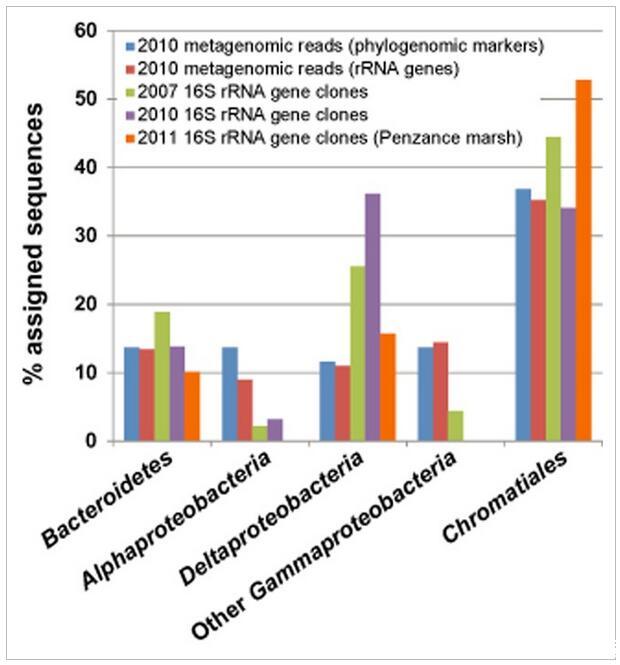
圖5
來自16S rDNA PCR擴增的克隆文庫與未組裝的250 bp雙端Illumina宏基因組讀數的細菌多樣性估計值的比較。宏基因組數據使用兩種不同的方法進行分析:通過Phylosift將最大似然讀數放置到系統發育保守的蛋白質編碼標記基因(“系統基因組標記”)上,或通過MG-RAST的M5RNA管道讀取與核糖體RNA基因的序列相似性。共有42 351個宏基因組讀數被歸類為細菌核糖體RNA序列,44 980個讀數被歸類為細菌系統發育標記基因。
使用共同組裝的Roche 454 Titanium和Illumina HiSeq序列數據對微生物多樣性進行了進一步的宏基因組分析。針對社區基因組學優化的策略(Peng等人,2012年)用于組裝7400個大于1千堿基(kb;N50 1.6 kb,最大重疊群64 kb)的支架。宏基因組支架的聚類(參見實驗程序)揭示了屬于PB-PSB1(20-25倍覆蓋)和PB-SRB1(15-20倍覆蓋)的兩個明確定義的箱。這些基因組箱的覆蓋深度對應于16S rRNA基因PCR數據中觀察到的PB-SRB1與PB-PSB1的豐度比(每1.1-1.7個PB-PSB1細胞1個PB-SRB1 RB)。這些基因組箱代表PB-SRB1和PB-PSB1的近乎完整的基因組,通過組裝序列長度、編碼特征的數量和對45個單拷貝系統發育標記基因的分析進行評估(參見支持信息,表S1)。在本研究中,我們將這些基因組的進一步分析重點放在與這些生物的硫基代謝相關的途徑上。
硫循環的宏基因組證據
為了評估漿果中硫循環生物的多樣性,我們將未組裝的宏基因組序列讀數映射到異化亞硫酸鹽還原酶基因(基因dsrAB,參見實驗程序)的系統發育,這些是氧化和還原異化硫代謝的廣泛使用的系統發育標記(洛伊等人,2008年)。大約33%與比對的dsrAB讀數序列無法通過這種方法進行分類,可能是因為它們源自系統發育無信息區域dsrAB基因的。大多數分類讀取被放置在Chromatiaceae(44%)或Desulfobulbaceae(13%,Supporting Information,Fig.S5)中。在這些家族中,大部分讀數被分配到Halochromatium salexigens(Chromatiaceae)和Desulfofustisglycolicus(Desulfobulbaceae),與顯性16S rRNA基因序列的系統發育隸屬關系一致。剩余的分類讀數(總比對讀數的10%)分布在Desulfobacteraceae和Desulfotomaculum中。
為了進一步表征粉紅色漿果硫氧化還原循環的潛力,我們從完整組裝的宏基因組數據集中確定了硫酸鹽還原和硫化物氧化的途徑。雖然在還原和氧化途徑中使用了許多相同的基因,但可以通過不同的序列和基因組背景清楚地區分同源物。通過基于獨立序列組成的分析,在與PB-PSB1分箱的支架上發現了所有鑒定的硫氧化基因,而硫還原基因位于PB-SRB1分箱中的支架上。PB-PSB1和PB-SRB1基因組中氧化和還原代謝途徑的詳細討論見支持信息S1、支持信息S2、表S2和圖S7-12
可溶性硫地球化學:物種形成、豐度和同位素組成
在缺氧過濾消毒的沼澤水中照亮的漿果微觀世界不會產生可通過Cline測定檢測到的硫化物(Cline,1969年),而是在兩天內迅速消耗了1 mM添加的硫化物(圖6 A)。為了研究聚集體中的硫形態和氧化還原環境,我們使用插入大漿果(直徑約0.5厘米)的金汞合金微電極進行了循環伏安法。從聚集體內部測量的伏安圖顯示電位為0.85 V(相對于Ag/AgCl)的峰值,隨著電極穿入和穿出漿果而變化。該測量對應于在內部漿果不同點13和41之間μM硫化物的濃度(圖6 B;還參見Supporting Information,Fig.S6)。使用克拉克型硫化物微電極(Unisense,Aarhus,Denmark)在其他大漿果內部進行的獨立分析表明存在5–20μM的H 2 S(135–540μM的總硫化物)。到通過循環伏安法可檢測到的其他電活性物質(例如O 2、Fe 3+、Fe 2+、Mn 2+和As 3+在漿果中沒有觀察、連四硫酸鹽、溶解/納米顆粒元素硫)。
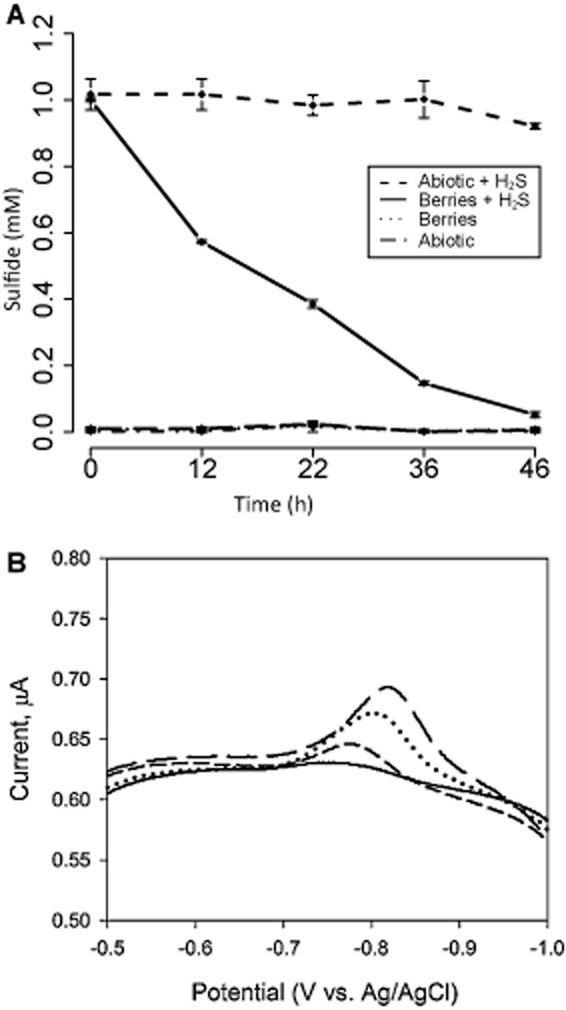
圖6
A.在50毫升過濾消毒的沼澤水中用50個小漿果進行微觀孵化時,通過Cline測定法測量硫化物濃度。孵育保持在14小時光照、10小時黑暗循環中。在漿果存在下的孵育過程中消耗了添加至1 mM濃度的硫化物(實線),而非生物對照顯示沒有變化(虛線)。沒有添加硫化物的微觀世界從接近0 mM的初始硫化物濃度沒有可檢測到的變化。誤差棒顯示三個生物重復孵化的標準偏差。
B.對直徑約0.5厘米的粉紅色漿果內的金汞合金電極進行伏安掃描顯示,當電極尖端穿透漿果時,在約-0.8 V(相對于Ag/AgCl)處出現與溶解硫化物相關的明顯峰。由于幾乎無法控制電極在漿果中的確切位置,因此數據并未量化硫化物的梯度,而是概述了整個聚合體中位置和相對值的變化。當電極在周圍的水中并且沒有穿透聚集體時,收集到沒有明顯峰的實線。
為了研究漿果中硫化物的同位素組成,將大的聚集體(直徑約0.5厘米)穿在24號銀線上并孵育過夜原位(圖7A)。可溶性硫化物以硫化銀的形式沉淀在焊絲上,在每個聚集體下方留下帶有金屬光澤的黑色光澤(圖7 B)。可見的硫化物積累開始于聚集體的表面下方,最暗的硫化物沉積在聚集體的中心,證實了來自微伏安法和硫化物微傳感器的硫化物報告。隨后使用IMS 7f-GEO扇形磁場二次離子質譜儀(SIMS;CAMECA,Gennevilliers,France)按照Fike及其同事(描述的方法分析了這種沉淀的硫化銀的豐度和同位素組成2009年)。沿著穿過兩個漿果的線的橫斷面顯示32 S計數(硫化物豐度)增加,δ減少34 S朝著聚集體中心(圖7 C)。在未發生可見硫化物沉積的情況下(即在漿果邊緣或漿果之間),32 S計數也非常低(圖7 C)。
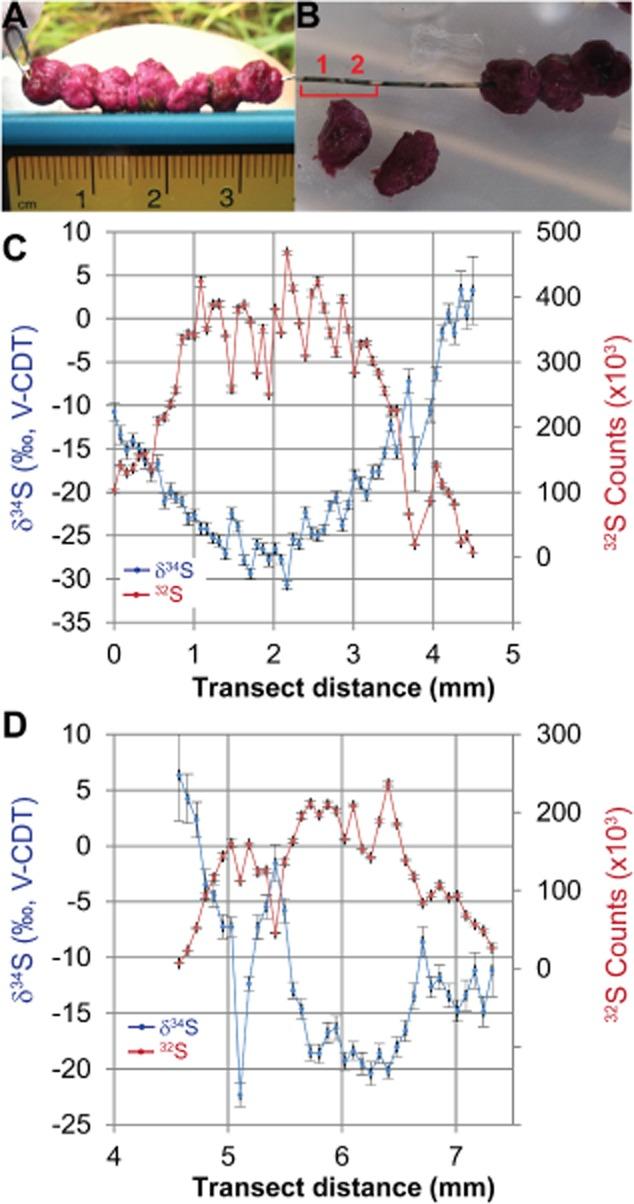
圖7
A.將大的粉紅色漿果穿在24號銀線上并孵育原位過夜。
B.漿果內產生的硫化物沉淀到金屬絲表面,形成一層AgS薄膜,可見漿果所在的黑色金屬光澤。SIMS7f離子微探針分析是在橫斷面(25μm光斑尺寸)中進行的,沿著導線穿過來自兩個不同漿果的AgS薄膜,這些漿果分別用紅色數字1和2標記,分別顯示在面板C和D中。
光盤。y軸上共同繪制的是的硫豐度(32 S計數,紅色)和δ34沿橫斷面(x軸)每個點S(藍色)。垂直誤差條代表的標準誤差(n每次測量=20個循環)。漿果1和漿果2之間32 S計數接近0的區域對應于導線上看不到深色硫化物膜的區域。圖7的SIMS數據也已在支持信息S2中提供。
的δ34沉積硫化物S范圍從聚集體邊緣的~6‰到聚集體中心的-31‰。相對于第一個漿果,沉積在橫斷面第二個漿果中心的硫化物顯示出較低的32 S計數和較少的同位素消耗(δ34 S為-20‰)(圖7 C)。潮間帶水體硫酸鹽δ34 S為+22‰;因此,這些“漿果內”同位素值表明同位素分餾范圍從~15‰到53‰。相比之下,在附近沉積物表面孵育但未穿透漿果(例如圖中最左側的環)的銀絲7Aδ34具有-16±2‰的S(平均值±SD,n=15)沿線均勻分布。
使用通過穩定同位素探測研究硫循環34 SO 4和nanoSIMS
為了跟蹤聯盟成員之間硫代謝和循環的活動,將新鮮收集的粉紅色漿果與下厭氧培養34富含S的硫酸鹽和富含13 C的碳酸氫鹽在光照和黑暗條件。,富集34培養4天后S在形態獨特的PB-PSB1細胞中的很明顯(圖8A和B)。的34 S/32這些細胞中S比率大約是未標記對照條件下觀察到的兩倍。發現該信號在甲醇中不穩定(圖甲醇8 C),是一種用于定量萃取元素硫的溶劑(Ferdelman等,1997)。富集34 S IN光亮和黑暗孵育(圖中記載8 C)。
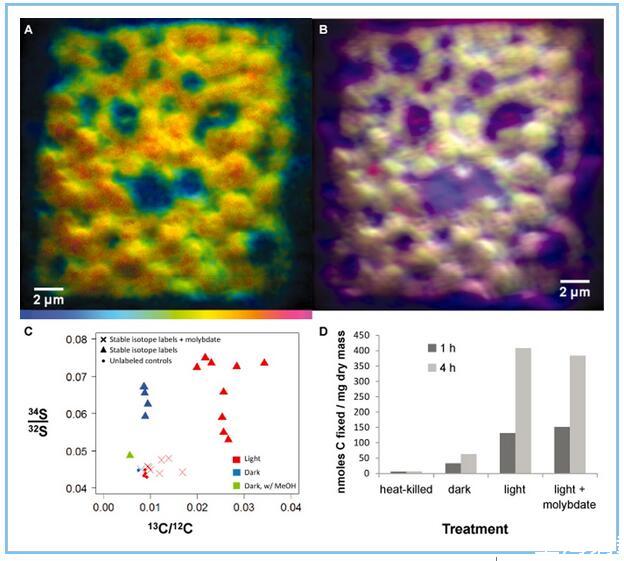
圖8
用同位素標記的粉紅色漿果的NanoSIMS分析34富含S的硫酸鹽和富含13 C的碳酸氫鹽。
A.色相-飽和度-強度圖像映射34 S:32 S比率。色標范圍從藍色(設置為在未標記的控制條件下觀察到的基線比率(0.044))到紅色,其中該比率相對于基線(0.08)增加了約2倍。顯示的圖像是來自聚集體外圍附近的粉紅色漿果生物量橫截面的復合八個連續30μm幀,分辨率為512×512像素。
B.來自疊加的初級離子信號的RGB合成圖像,顯示了形態獨特的紫硫細菌細胞的細胞排列。疊加的初級離子圖像是12 C離子(藍色)、12 C 14 N(綠色)和32 S(紅色)。提供的每個質量的基礎離子圖像支持信息S2中。
C.NanoSIMS計算的穩定同位素標記的平均摻入13 C-碳酸氫鹽(x軸)和34 S-硫酸鹽(y軸)在黑暗(藍色)或12小時光/暗循環(紅色)中孵育4天的在活性硫酸鹽呼吸(三角形)或鉬酸鈉(X)抑制SRB的條件下,與未標記的對照孵育(點)進行比較。用甲醇(綠色三角形)對同位素標記的深色孵育漿果進行后處理。繪制的值是來自不同區域的15×15μm(淺色)或30×30μm(深色)柵格的平均值,總共七個聚合體(每個條件一個)。
D.將14 C碳酸氫鹽摻入與1 mM硫化物一起孵育1小時或4小時的五個漿果中的酸穩定產品中。對熱滅活的聚集體、暗平衡聚集體和在光照下添加和不添加10 mM鉬酸鹽的聚集體進行孵育。
積累34 S同位素標記的似乎取決于聚集體中硫酸鹽還原生物的活性;此外鉬酸鈉(NaMoO的4),硫酸還原的抑制劑(Oremland和卡彭,1988),阻斷的摻入34 S-富集標簽(圖8 C)。發音的34 S-富集(1100±200‰為三次重復溫育)在散裝漿果生物質中觀察到從孵育用34 S-富集硫酸鹽,如通過元素分析儀同位素比率質譜儀(確定支持信息,圖S13)。未標記的對照和鉬酸鹽處理的散裝生物質的同位素組成與直接從沼澤中取樣的天然漿果相當(δ34 S值范圍從-15‰到-24‰)。
碳固定是光依賴性的,如通過nanoSIMS測量所評估的13 C摻入(圖8 C)和放射性標記(14 C)碳酸氫鹽孵育測定(圖8 D)的,表明PB-PSB1作為主要生產者在聚合中。鉬酸孵育不影響在短期溫育(1-4 1H)碳的光依賴性固定,表明PB-PSB1不是直接由鉬酸(圖抑制8 d)。在4天的溫育,鉬酸發現以降低聚集體碳固定在光,這表明較長的持續時間的治療鉬酸強加限制硫化物,從而降低PB-PSB1的初級生產力(圖8 C)。
《鹽堿地沼澤中的光養粉紅色貝類的微量硫循環》——概括 、介紹
