熱線:021-66110819,13564362870
Email:info@vizai.cn
熱線:021-66110819,13564362870
Email:info@vizai.cn
2.1、物種和生物材料
本研究中調查的物種的選擇主要是因為它們的生態重要性或經濟相關性:它們沿著 Sungo Bay 海岸線豐富,雖然不跨越其門的真正分類范圍,但很好地代表了當前的群落組合。 Sungo Bay 大型藻類群落本身可與全球其他海岸線相媲美。 例如,Macrocystis pyrifera 最初是從墨西哥進口的,現在在水產養殖中進行商業種植。 同樣,糖精最初于 1970 年代從日本引進,此后成為中國最重要的商業種植海藻。
2.2、樣品采集
M. pyrifera 的成熟孢子體最初于 1978 年在墨西哥圣托馬斯(MST,31°330N,116°240W)收集。本研究中使用的譜系的雄性和雌性配子體克隆被分離并保存在黃海漁業條件包括溫度、鹽度和 TA 的監測,如對室內中層世界所述,以分析海水化學的變化(圖 S6)。 如上所述,通過量熱法測定營養素。 為了避免假復制,每個 pCO2 水平設置了六個籠子; 然而,由于物流限制(這些類型的實驗的一個常見問題,康沃爾和赫德,2015 年也強調了這一點),我們無法在單獨的網箱中種植每個物種。 每個物種的 20 個重復在每個網箱中生長。 雖然平均溫度 (9.38 ? 2.56°C) 與室內中層的溫度相當,但這里的溫度也隨著時間的推移而升高,因為實驗是在晚春月份進行的。 此外,室外和室內設置之間的 pCO2 平均值和方差具有可比性,而室外中層環境中營養物質和光強度的變化大于室內設置。 然而,平均光照強度和日長與室內中層環境相當。 為了將此數據集與室內中觀的數據集進行比較,還以 7 天的間隔記錄了 28 天的生長數據。
2.4、生長、光合作用、呼吸作用的測定
藻類比生長率 (SGR) 的確定和計算如下:
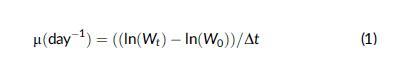
式中 W0 和 Wt 為培養期開始和結束時的鮮重; Dt 是采樣間隔之間的天數。
使用描述單峰曲線的經驗增長模型對響應 pCO2 變化的增長率變化進行建模
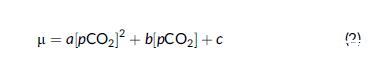
其中半飽和常數為 Km = -b/2a,最大比增長率為 lmax = (4ac - b2)/4a。 常數 a、b 和 c 是要為單峰增長方程確定的參數。
藻類凈光合作用 (NP, lmol g?1 新鮮 hr?1) 記錄在中午時我們預計它們是光飽和的(從 12:00 到 14:00)。 為了進行測量,將藻類樣品從中間宇宙中取出并放入密封的玻璃室中,玻璃室中充滿了來自每個水箱中具有適當碳酸鹽化學成分的海水。 使用氧電極 (Unisense) 原位測定氧濃度的變化,然后用于計算 NP。 在每個采樣時間的午夜(從 24:00 到 02:00)也使用相同的方法監測呼吸(RE,lmol g?1 新鮮 hr?1)。
2.5、統計分析
2.5.1 、實驗室實驗
由于在 6 天的時間里對裝有生物的罐子進行了重復采樣,以進行實驗室中最短的實驗,而在中宇宙中進行較長時間的實驗則需要 28 天,因此使用了 R 包 NLME(版本 3.1-128)中的混合效應模型分析 pCO2 對生理機能的影響。 生長速率被用作要解釋的實驗變量。 時間和重復被用作隨機效應(因為所有測量都以我們無法解開的方式與第一次測量相關)。 然后我們添加了“門”(即紅藻門、綠藻門或褐藻門)、“生命”(多年生與一年生,因為樣本在整個門中分布不均,并且褐藻不是嚴格認可的分類群),或“策略”(開花形成的“r 戰略家”與非開花的“K 戰略家”)作為一個固定效應,另一個固定效應是“二氧化碳水平”。 為了解決將不同物種分成更大組的可能注意事項,還測試了“物種”對自身的影響以及嵌套在更大組中的影響(例如,“門”和“策略”)。 然后分別測試“Phyla”、“life”、“strategy”和“species”,每個都與 pCO2 相互作用,并根據最小的 AICc 分數(Akaike Information Criterion for small sample size)選擇最簡約的模型進行報告)。 當 AICc 分數變化小于兩個單位時,通過 aictab 函數(AICCMODAVG 2.10 版)進行模型平均。
有關物種及其相關策略的完整列表,請參見表 S4。 在 GLHT 包(版本 1.46)中的“multcomp”函數中進行了成對事后測試。 混合模型的選擇,而不是方差分析,也說明了數據的不平衡設計(例如,“門”和“策略”中的子集數量不同)。 AICc 表在 SI 中可用(表 S5-S7)。 作者將根據合理要求提供完整的 R 代碼。
2.5.2 、室內外中觀
在這里,響應變量是生長速率、光合作用和呼吸速率。 時間再次被用作隨機效應,如 aquariaID 或 netcageID(即生物復制)。 然后對每個響應變量按上文所述進行混合效應模型。 我們使用與上述相同的固定和隨機效應來分析實驗室實驗,但還包括對其他非生物參數(例如,營養濃度、溫度)的測試,以測試另一個參數是否在推動物種反應或與變化顯著相互作用在 pCO2 中。
僅對于室內中層世界,為了將生理參數(光合作用和呼吸)的變化量與生長速率的變化量相關聯,我們計算了 pCO2 每次變化時每個性狀的倍數變化,即所有值在將高 pCO2 水平與 400 latm pCO2 的“對照”水平進行比較,并計算相對于該水平的變化。 然后通過線性回歸檢查生長變化與光合作用或呼吸變化之間的關系。
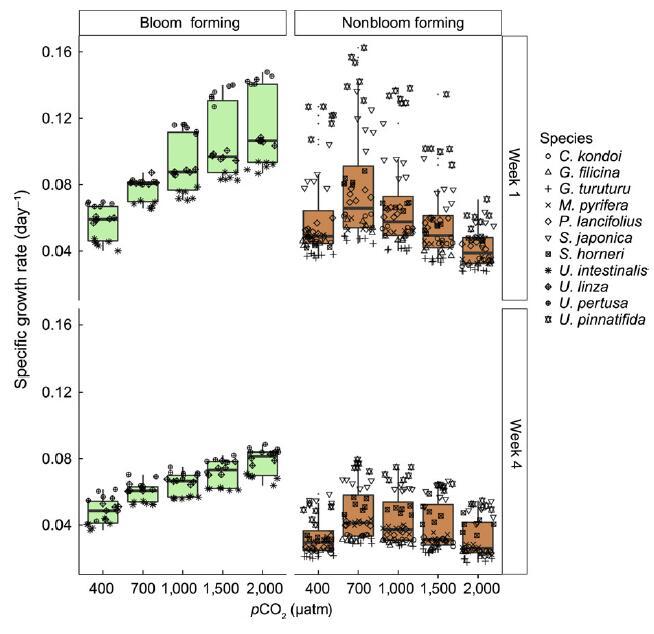
圖 2 生長響應曲線 成花(淡綠色)和非成花 (淺棕色)物種響應 在開始時升高 pCO2(一周 1,上圖)和結束(第 4 周,下圖) 面板)的室內潛伏期 中觀。 就像在實驗室里 治療,生長率響應 成花過程中 pCO2 水平升高 物種(雖然增加 增長不像過去那樣呈指數增長 實驗室環境),而非開花 物種表現出單峰 響應,增長達到峰值 700 和 1,000 latm pCO2 (F2,30 = 64.84, p < .0001)。 增長率下降 對所有場景中的所有物種都有顯著影響。 箱線圖按標準顯示,帶有 中位數作為粗體帶,和胡須 擴展到第一個四分位距。 覆蓋箱線圖的形狀表明 單個物種,每個物種的 n 為 每個 pCO2 水平 = 6。 [顏色數字可以 在 wileyonlinelibrary.com 上查看]
同樣對于室內中觀,進行了主坐標分析 (PCoA)(包“VEGAN”2.42),以使用獲得的 Km 和 lmax、光合作用和呼吸的結果分析對 pCO2 變化的生理反應的物種特異性差異。 所有主坐標都用于計算差異矩陣,然后測試它們對方差的貢獻。 選擇了解釋大部分(這里接近 100%)整體變化的兩個變量進行顯示。 為了比較室外和室內中觀之間的結果,“位置”被添加為一個額外的固定因素,全球模型包括 pCO2、物種(或策略)、時間和位置之間的完整相互作用。 然后使用“dregde”函數(MUMIN 版本 1.15.6)研究該模型,并選擇具有最小可能 AICc 分數的模型進行進一步討論和事后測試。
在環境復雜性海洋環境中馴化的藍藻和海藻能提高海洋中PCO2分壓總保有量——摘要、介紹
在環境復雜性海洋環境中馴化的藍藻和海藻能提高海洋中PCO2分壓總保有量——材料和方法
在環境復雜性海洋環境中馴化的藍藻和海藻能提高海洋中PCO2分壓總保有量——結果
在環境復雜性海洋環境中馴化的藍藻和海藻能提高海洋中PCO2分壓總保有量——討論、致謝!
 相關新聞
相關新聞