熱線:021-66110819,13564362870
Email:info@vizai.cn
熱線:021-66110819,13564362870
Email:info@vizai.cn
研究簡介:富含黑素體的偏振上皮細胞的視網膜色素上皮(RPE)位于視網膜感光層和布魯赫膜之間,靠近脈絡膜。RPE損傷與退行性視網膜疾病的發展有關,例如糖尿病視網膜病變(DR)和年齡相關性黃斑變性(AMD),這兩種疾病都是導致失明的主要原因。光感受器與RPE、布魯赫膜和脈絡膜的相互作用調節11-cis視網膜和光感受器外節(OS)的更新、神經視網膜和循環之間的物質交換,并形成外節血視網膜屏障(OBRB)。視桿細胞OS是一種參與視覺信號轉導的修飾纖毛。特別是該操作系統包含大約2000個堆疊的膜盤,它們經歷著受光調節的持續更新過程。當RPE不斷吞噬OS尖端盤時,新的盤在纖毛基部形成。OS吞噬作用涉及αvβ5整合素受體、粘著斑激酶(FAK)和Mer酪氨酸激酶(MerTK)。椎間盤的形成仍不完全清楚,它涉及纖毛旁邊外翻形成囊的基底膜的合成,以及通過鞭毛內轉運蛋白(IFT)摻入內部節段合成的椎間盤蛋白。視桿細胞的凋亡比視錐細胞更容易受到損傷,進而導致視錐細胞損失,損害高敏銳度視力。視桿細胞分泌一種營養錐活力因子,這種因子是在截短的硫氧還蛋白樣蛋白中發現的。
為了闡明DR的發病機制,了解RPE和視桿OS之間的相互作用似乎至關重要,考慮到為了維持光轉導的能量需求,OS表現出自己的氧化代謝。由于還沒有研究調查這一主題,本研究工作旨在評估OS和RPE之間是否存在串擾,從而導致外視網膜累積氧化損傷。因此研究人員研究了未氧化(UOx)或氧化(Ox)rod OS吞噬作用對在正常(NG)或高葡萄糖(HG)條件下生長的ARPE-19細胞的影響。
Unisense微呼吸系統的應用
在37°C的密閉室中通過微呼吸氧電極(Unisense Microrespiration,Unisense A/S)評估耗氧率(OCR)。在實驗中,將2×105 ARPE-19細胞重新懸浮在磷酸鹽緩沖液(PBS)中,并用0.03 mg/mL毛地黃皂苷透化1分鐘。總共添加10 mM丙酮酸加5 mM蘋果酸或20 mM琥珀酸,分別刺激由復合物I、III和IV或復合物II、III和IV組成的呼吸途徑。這兩種情況下,都添加了0.1 mM ADP。對于桿OS,使用50μg總蛋白,并使用0.1mM NADH和0.1mM ADP作為呼吸底物。
實驗結果
數據表明RPE和OS光感受器之間存在串擾,這在DR發病機制中起決定性作用。這些數據還支持了新的發現,即DR的首要推動因素是對視網膜外層神經元的氧化損傷。后者反過來會對RPE造成氧化損傷,并最終對血管系統造成氧化損傷。數據還表明單獨的ARPE-19細胞可能并不是研究外視網膜氧化應激作用的詳盡模型。研究人員認為評估幾種細胞類型之間的相互作用是必要的。目前的實驗模型可以突出體內病理條件下可能發生的潛在現象,為未來DR的預防性干預開辟新的視野。然而有必要在動物模型中進行進一步的體內實驗,以證實關于高血糖條件下RPE和rod OS之間氧化應激產生中的串擾的體外數據。
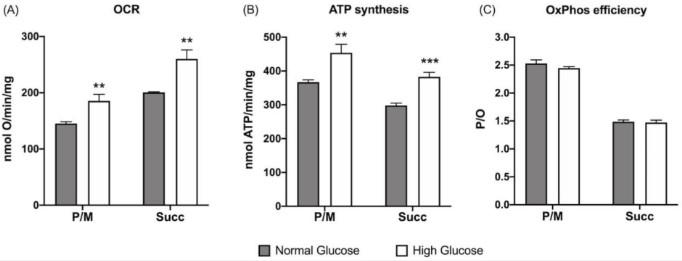
圖1、生長培養基中葡萄糖濃度對ARPE-19細胞有氧代謝的影響:(A)耗氧率(OCR);(B)有氧ATP合成;(C)P/O值作為OxPhos效率標記。在存在丙酮酸加蘋果酸(P/M)或琥珀酸(Succ)的情況下評估了有氧代謝,以分別刺激復合物I、III和IV以及復合物II、III和IV的途徑。灰色柱代表在5.6 mM葡萄糖(正常葡萄糖,對應于通常用于生長這些細胞的葡萄糖濃度)下生長的ARPE-19細胞獲得的數據,白色柱代表在高葡萄糖培養基(25 mM)中生長的相同樣品。
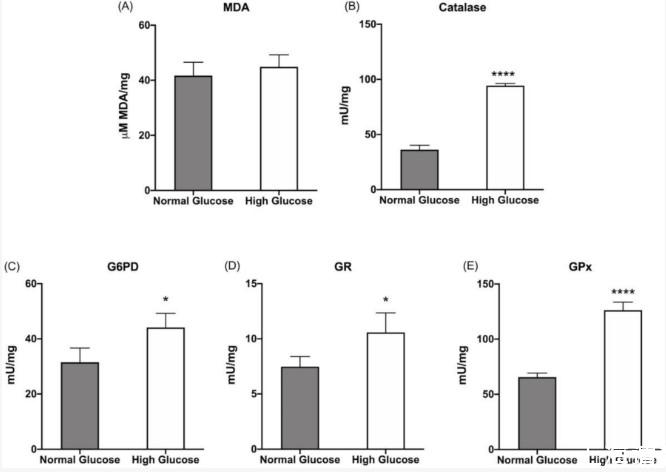
圖2、在正常或高葡萄糖培養基中生長的ARPE-19細胞中的脂質過氧化和抗氧化防御:(A)丙二醛(MDA)濃度作為脂質過氧化的標記;(B)過氧化氫酶活性;(C)葡萄糖6-磷酸脫氫酶(G6PD)活性;(D)谷胱甘肽還原酶(GR)活性;(E)谷胱甘肽過氧化物酶(GPx)活性。所有測試的酶都參與細胞抗氧化防御。灰色柱代表在5.6 mM葡萄糖(正常葡萄糖,對應于通常用于生長這些細胞的葡萄糖濃度)下生長的ARPE-19細胞獲得的數據,白色柱代表在高葡萄糖培養基(25 mM)中生長的相同樣品。
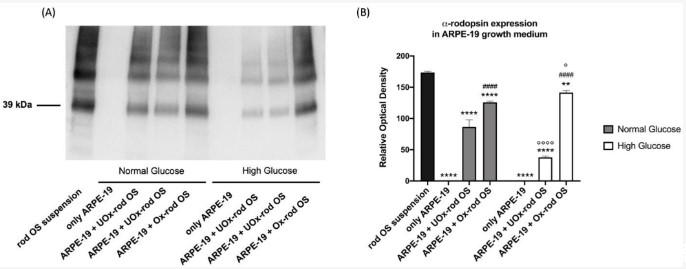
圖3、ARPE-19細胞生長培養基中的視紫紅質數量作為細胞吞噬未氧化或氧化的視桿細胞能力的標志。(A)與未氧化(UOx)或氧化(Ox)視桿OS孵育5.5小時后ARPE-19細胞生長培養基中的視紫紅質信號。補充圖S2報告了整個WB信號,包括分子量(MW)標記。(B)視紫紅質單體信號(39 kDa)的光密度分析。黑色柱代表孵育前rod OS懸浮液中視紫紅質的濃度;灰色和白色柱分別代表NG和HG條件下的視紫紅質濃度。數據表示為平均值±SD,代表四次獨立重復(n=4)。
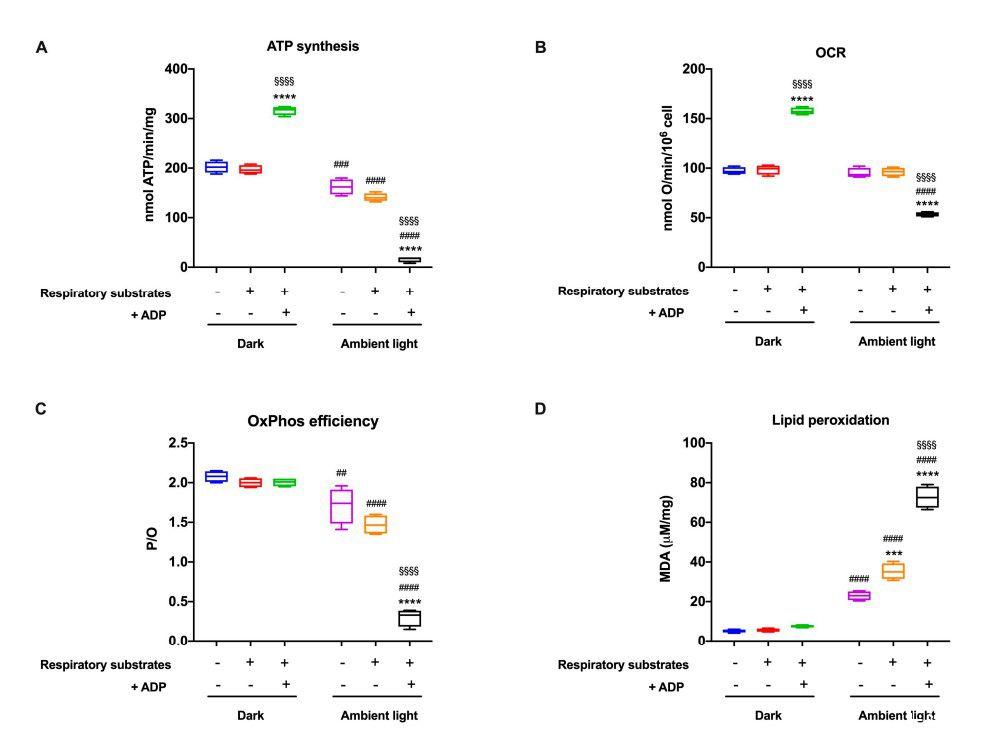
圖4、ATP合成、耗氧率、氧磷效率和脂質過氧化。圖所示的所有數據均已獲得從與呼吸底物(0.6 mM)預孵育或未孵育的牛棒狀OSs中提取NADH和20 mM琥珀酸鹽/或0.1 mM ADP,并在黑暗中維持(A)通過F1Fo-ATP合成ATP。合成酶和(B)耗氧量(OCR);在兩個實驗中都是0.1 mM加入NADH誘導OCR和ATP合成。(C)P/O值,OxPhos效率的標志,計算為合成ATP與消耗氧氣的比值。(D)MDA含量作為脂質過氧化標志物。數據具有四個獨立重復的代表性(n=4)。
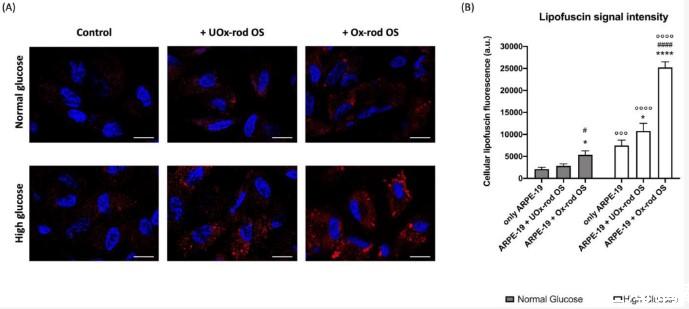
圖5、在正常或高葡萄糖培養基中生長并與UOx-或Ox-rod OS一起孵育的ARPE-19細胞中脂褐素的積累。(A)代表性共焦圖像,報告在正常葡萄糖(NG)和高葡萄糖(HG)條件下生長并與UOx-或Ox-rod OS一起孵育的ARPE-19細胞中細胞質脂褐質積累(紅色信號)。藍色信號對應于用DAPI染色的細胞核。條形刻度對應于10μm。(B)脂褐質熒光信號的強度。灰色柱代表在NG條件下生長的ARPE-19細胞獲得的數據,白色柱代表在HG條件下生長的相同樣品。
結論與展望
視網膜色素上皮(RPE)功能障礙與多種以視網膜變性為特征的疾病有關,例如糖尿病性視網膜病(DR)。然而外層視網膜神經元也參與損傷觸發。研究人員評估了RPE和光感受器之間在引發和維持RPE氧化損傷方面可能存在的串擾。使用ARPE-19細胞作為人類RPE模型,在正常(NG,5.6 mM)或高葡萄糖(HG,25 mM)和未氧化(UOx)或氧化(Ox)哺乳動物視網膜桿外節中生長。ARPE-19細胞在NG和HG環境中都能有效吞噬rod OS。然而在HG中,與未處理的細胞和UOx-rod-OS處理的細胞相比,用Ox-rod OS處理的ARPE-19細胞積累了MDA和脂褐素,并表現出LC3、GRP78和caspase 8表達的改變。數據表明早期氧化損傷可能源自光感受器,隨后延伸至RPE,這為視網膜變性僅取決于RPE氧化還原變化的觀點提供了新的視角。
